The 10-year average annual cycles of D and B calls through time indicates that whales arrive at the SCR feeding grounds in May and depart in November, remaining at the feeding grounds an average of 8.4 months (Fig. 1, Supplementary Table 1, Supplementary Fig. 3). The timings of D call onset and cessation displayed greater variability than B call onset and cessation (Fig. 1), suggesting that D calls may be influenced more by external forces than B calls. There was no significant relationship between the duration of overlap of D and B calls in the same year, or between the timings of B call cessation and D call onset the following year. The onset, cessation, and duration of both D and B calls displayed interannual variability, suggesting that the timing of these calls, and, by inference, blue whale arrival and departure, was not associated with photoperiod, as has been documented for many terrestrial birds and mammals25. Instead, blue whales must use other cues to detect interannual variability and determine when to migrate to and from their feeding grounds.
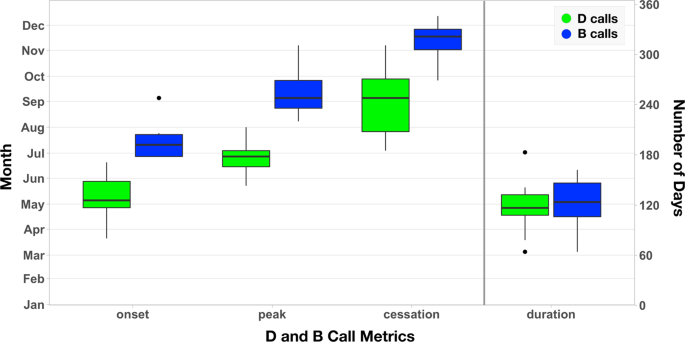
Summarized D and B call metrics for date of onset, peak, cessation, and duration (number of days). Each whisker boxplot displays the median, first and third quartiles (25th and 75th percentiles), upper and lower whiskers (1.5 times the inter-quartile range), and outliers (black circles).
Arrival time at the feeding grounds is correlated with sea surface temperature anomalies from the previous feeding season
Eight-day mean sea surface temperature (SST) anomalies integrated over May to November of the previous feeding season (the average time whales are in the SCR) were correlated with the timing of the onset of D calls (i.e., blue whale arrival) in the SCR the following year (multiple regression partial R2 = 0.78, p < 0.01; Fig. 2). Specifically, when the previous feeding season was colder, D call onset (i.e., arrival) in the SCR was earlier the following year, and it was later following warmer years.
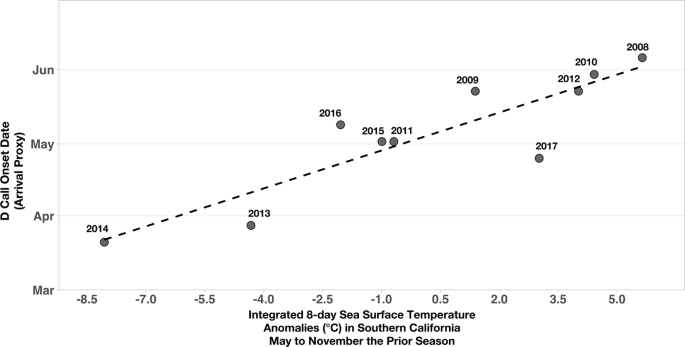
D call onset at the Southern California feeding grounds compared to integrated 8-day sea surface temperature anomalies the prior feeding season. Annual (2008–2017) onset of D calls (our proxy for blue whale arrival at the feeding grounds) correlated with integrated eight-day sea surface temperature anomalies (°C) in Southern California from the prior feeding season (May–November; multiple regression partial R2 = 0.78, p < 0.01).
Temperature is a migratory cue used by many terrestrial taxa—from insects to birds to ungulates6,26. In the marine realm, SST is similarly important for highly migratory species, including leatherback sea turtles (Dermochelys coriacea)27 and flounder (Platichthys flesus)28. Nishiwaki29 was the first to suggest that changes in SST might influence the timing of humpback whale (Megaptera novaeangliae) arrival on their wintering grounds. Visser et al.30 tied the timing of peak baleen whale abundance in the Azores to rising SSTs following the spring bloom, and Tsujii et al.31 found that water temperature was a good predictor of fin whale (Balaenoptera physalus) arrival and departure in the southern Chukchi Sea. Our study demonstrates how oceanographic conditions influence migration timing, but also suggests the use of memory.
The idea that memory of past conditions combined with resource tracking allows animals to modify the time, speed, and direction of migration movement has been documented in terrestrial mammals32 and birds, including demonstrating the memory of high-quality foraging locations for at least 12 months33. Among the limited number of studies of the timing of whale migration, to our knowledge, only one examined the role of memory. Abrahms et al.34 found that tagged blue whale latitudinal migratory movements correlated with a 10-year average spring bloom (via chlorophyll-a peaks) and hypothesized a long-term memory of the location of highly productive foraging sites. Our findings, and the fact that we could detect no relationship between D call onset and any environmental variable on the breeding grounds prior to arrival at the feeding grounds, support the hypothesis of memory use in migratory timing. By integrating the SST anomaly signal at the feeding grounds from the prior feeding season, whales may forecast future conditions and adjust their arrival timing the following year.
Annual sea surface temperature anomalies from the previous season are correlated with krill biomass at the feeding grounds
Colder annual SST anomalies in the SCR the previous feeding season were associated with greater krill biomass the next year, while warmer annual SST anomalies were associated with lower krill biomass (R2 = 0.34, p = 0.06; Fig. 3). Previous studies have established that associations between colder water and greater zooplankton biomass result from the upwelling and advection of cold, nutrient-rich water and subsequent primary production1. Greater krill biomass in the SCR was also associated with an earlier onset of D calls, when whales arrived at their feeding grounds (multiple regression partial R2 = 0.52, p = 0.04; Fig. 4). The relationship between SST anomalies, krill biomass, and D call onset suggest that in addition to anticipating future conditions based on the prior year’s conditions, whales could profitably use SST as a proxy for krill biomass. Thus, whales could optimize arrival time at the feeding grounds to take advantage of abundant prey (cold years, early arrival) or limit their effort in the area when they expect prey to be impoverished (warm years, late arrival).
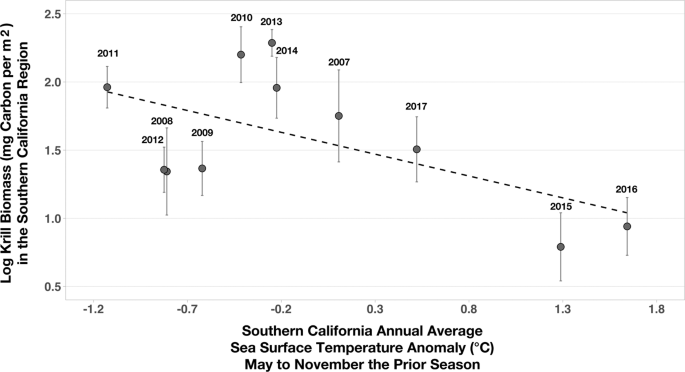
Krill biomass compared to annual sea surface temperature anomalies in Southern California the prior feeding season. Annual (2008–2017) spring biomass (log transformed; mg carbon per m2 with standard error bars) of adult and juvenile Euphausia pacifica and Thysanoessa spinifera correlated with annual sea surface temperature anomalies (°C) in Southern California from the prior feeding season (May-November; R2 = 0.43, p = 0.07).
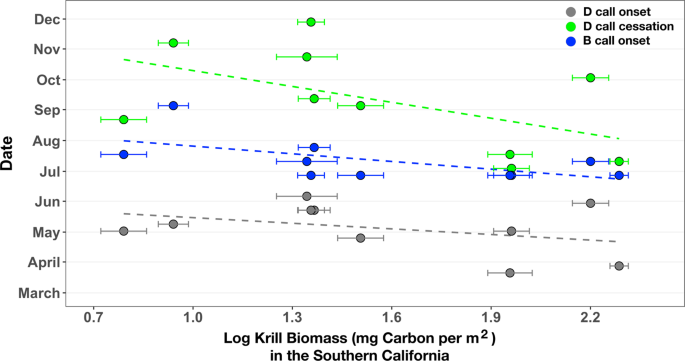
Krill biomass in the Southern California Region compared to D and B call onset and D call cessation. Annual (2008–2017) spring biomass (log transformed mg carbon per m2 with standard error bars) of adult and juvenile Euphausia pacifica and Thysanoessa spinifera correlated with the onset of D calls (multiple regression partial R2 = 0.52, p = 0.04), cessation of D calls (R2 = 0.28, p = 0.10), and onset of B calls (R2 = 0.36, p = 0.07).
Analyses of blue whale scat collected in the SCR have shown that whales preferentially target T. spinifera20, which has a greater lipid content than E. pacifica35, though both euphausiid species compose their diet and are dominant in the cold waters of the SCR36. Zooplankton in cold, high-latitude waters have higher energy contents and lipid stores than zooplankton at lower latitudes37. Given the potential for a greater caloric value from cold-water compared to warm-water euphausiid prey, there may be a significant feeding advantage for whales to arrive on their feeding grounds earlier when it is colder, and when lipid-rich krill is predictably more abundant.
During warmer years, it is possible that whales delay departure from the CRD to opportunistically feed locally on smaller, less energy-rich krill such as Euphausia eximia, Euphausia gibboides, Euphausia distinguenda, Nematoscelis gracilis, Nematobrachion flexipes, and Nyctiphanes simplex before migrating to the SCR38,39. However, while some data suggest that whales may feed in the CRD15, there is no evidence that the bulk of this population remains year round or that the euphausiid species available at blue whale breeding grounds40 could support the energetic demands of the population. Another possibility, suggested from analyses of visual, acoustic, and satellite tagging data15,40,41, is that in warm years blue whales leave the CRD, but stop in the Gulf of California and along Baja California Peninsula, where they feed opportunistically on the subtropical euphausiid Nyctiphanes simplex on their way to a relatively less-productive SCR.
Krill biomass mediated the transition to reproductive-related calling behavior at the feeding grounds
Higher krill biomass in the SCR was associated with earlier D call (multiple regression partial R2 = 0.52, p = 0.04; Fig. 4) and B call (R2 = 0.36, p = 0.07; Fig. 4) onset there, as well as earlier D call cessation (R2 = 0.28, p = 0.10; Fig. 4). In years when whales had access to greater-than-average krill biomass, they ceased D calls sooner and started to produce B calls sooner. Also, in years with greater krill biomass, the duration of D calling was shorter, though not significantly (R2 = 0.23, p = 0.16). The production of song by male humpback whales has been studied extensively, including on feeding grounds42,43, and proposed hypotheses regarding the purpose of these songs include intersexual and intrasexual functions44. While the production of B calls by male blue whales is likely associated with reproduction, their calling behavior is complex and, like humpback whale song, the precise functions are unknown22. However, our discovery of the link between higher krill biomass at the feeding grounds and blue whales’ earlier transition to reproductive-related calling behavior (i.e., onset of B calls) reinforces the hypothesized importance of the connections among migration timing, prey quality, and reproductive-related behavior25,45.
The time of arrival of blue whales to the feeding grounds showed long-term trends
The onset of D calls (our proxy for blue whale arrival at the SCR feeding grounds), showed a long-term trend of earlier onset over the 10-year period (multiple regression partial R2 = 0.53, p = 0.04), shifting more than one month (42 days) from June to April. Across the same 10-year period, mean annual SST in the SCR increased by 1 °C (R2 = 0.55, p = 0.01; Fig. 5b). We hypothesize that the decadal warming trends are driving the whales to arrive at the SCR feeding grounds earlier. This has led to whales spending more and more time at the SCR feeding grounds.
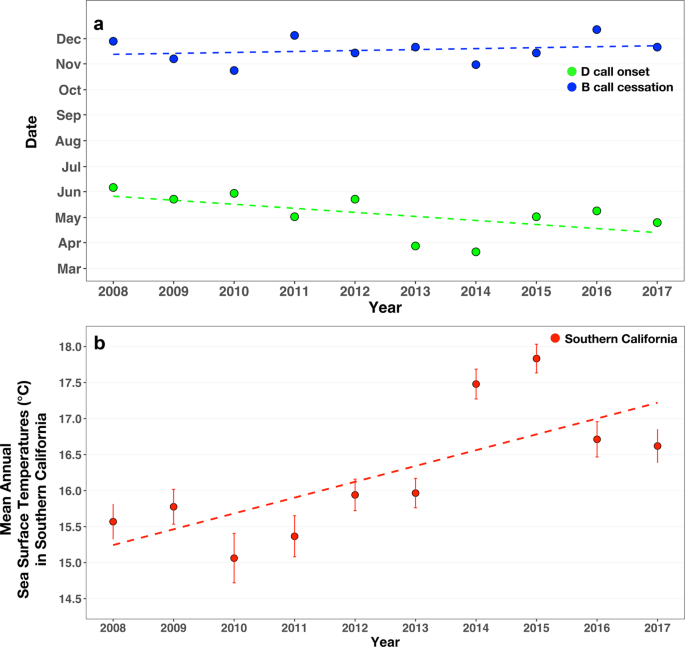
Long-term trends in D call onset, B call cessation, and annual average sea surface temperatures. (a) D call onset date (i.e., arrival at the feeding grounds) shifted significantly earlier across the 10-year study period (2008–2017; multiple regression partial R2 = 0.53, p = 0.04) while there was no significant shift in b call cessation (i.e., departure from the feeding grounds) (R2 = 0.05, p = 0.53). (b) Across the same time period, there was a significant increase in average annual sea surface temperatures (°C with standard error bars) in Southern California (R2 = 0.55, p = 0.01).
Long-term change in migration timing has been demonstrated in amphibians, birds, insects, fish, marine invertebrates, marine zooplankton, and mammals6,46. There are few studies of long-term temporal changes in migration timing for highly migratory aquatic animals; however, the continued collection of photographic identification, passive acoustic monitoring, and satellite tag data, is revealing early arrival trends in other whale species. Across a 27-year period, fin and humpback whales arrived one month earlier on their feeding grounds in the Gulf of St. Lawrence in the North Atlantic Ocean. Because this timing shift was significantly correlated with increased sea-surface temperature and decreased sea-ice formation, it was hypothesized that the shift in arrival allowed whales to track changes in the timing of the spring bloom47. A similar study using telemetry and acoustic data found that over a 22-year period, beluga whales (Delphinapterus leucas) in the Eastern Beaufort Sea departed later in years with delayed sea-ice freeze-up, which likely enhanced productivity and zooplankton advection48.
Our findings show that blue whales have altered their timing of migration in the CCE of the United States. We hypothesize that as the waters in the SCR and CRD are warming, the quality and quantity of krill biomass are changing, removing any previous advantage of remaining longer in the CRD or along the coast of Baja to feed on smaller, less energy-rich species of krill. Krill biomass in the SCB also shows a long-term increase49, suggesting an advantage for blue whales spending more time at the feeding grounds off the California coast. In addition to changes in krill biomass, previous studies have documented krill range contractions, and range shifts coincident with physical oceanographic changes50, which may further influence blue whale migration behavior.
These long-term adjustments to changes in prey distribution and availability may result in whales following their prey poleward, remaining on feeding grounds longer, or suspending migration. Bioenergetic models indicate that increases in travel time resulting from poleward expansion increase the overall energetic cost of migration and reduce the time available for feeding, reproduction, and calving51. A longer migration or feeding period may result in a decreased frequency of migration, especially if the cost of migration becomes too high51,52. Any adjustments by the whales to track changes in prey distribution and biomass may also increase their spatial overlap with anthropogenic threats, further threatening this already endangered species. For example, in the case of this population, because of the high volume of ship traffic in the SCR, increased residence time could increase the whales’ lifetime risk of being struck by a ship53.
The time of departure of blue whales from the feeding grounds shows long-term stability
The cessation of B calls, our proxy for blue whale departure from the feeding grounds did not change across the 10-year study period (R2 = 0.05, p = 0.53, Fig. 5a). In other mammals, migration phenology has been shown to be influenced by a combination of external biotic and abiotic cues, as well as by endogenous biological clocks regulating the physiological and morphological changes necessary for these behaviors6. Hormones linked to migration timing include melatonin6, adipose and thyroid hormones, and gonadal steroids54. Although virtually unstudied in the context of migration for marine mammals, these hormones likely play a role in their transition from feeding to reproductive-related calling, possibly triggering whale migration back to their breeding grounds. Because the cessation of B calls was not related to any environmental indices or krill biomass at the feeding grounds, we hypothesize that the cessation of B calls, our proxy for departure from the feeding grounds, may be partially regulated by seasonal fluctuations in hormone levels.
Leptin, which has been studied in bowhead whales (Balaena mysticetus) and beluga whales55 is a satiety hormone that is released as adipose tissue increases, signaling to the reproductive system that sufficient fuel reserves have been stored to support reproduction25 and stimulating ovulation in female mammals56. Additional evidence of the influence of hormones comes from the analysis of cross-sections of baleen from a stranded male blue whale, which displayed regularly spaced areas of high testosterone peaks57. Although the age of the whale was unknown, the cycles mirrored annual cycles of testosterone measured in the baleen of a bowhead whale and North Atlantic right whale (Eubalaena glacialis) of known ages57. Seasonal fluctuations in testosterone have also been measured from blubber samples of male humpback whales with mean testosterone peaks between November and January58,59. The similarity in annual testosterone cycles for these baleen whale species and similarities between humpback and blue whale reproduction support the idea that hormones play a role in migration phenology, especially triggering departure back to the CRD breeding grounds.
Source: Ecology - nature.com
