Systematic paleontology
Thalattosauria Merriam, 1904
Thalattosauroidea Nopsca, 1928
Gunakadeit gen. nov.
Gunakadeit joseeae sp. nov.
Etymology
The generic name is derived from Gunakadeit, (pronounced Goo-na’-ka-date), a sea monster of the Tlingit culture. The specific ending honors Joseé Michelle DeWaelheyns, mother of the discoverer.
Holotype
UAMES 23258 (Figs. 2, 3, 4, 5, 6 and 7) a nearly complete skeleton including the skull and most of the axial and appendicular skeleton; missing the distal two-thirds of the tail and the distal portions of the limbs. The holotype is housed in the Earth Sciences Collection at the University of Alaska Museum (UAMES).

Gunakadeit joseeae gen. et sp. nov. Holotype specimen UAMES 23258 in right lateral view.
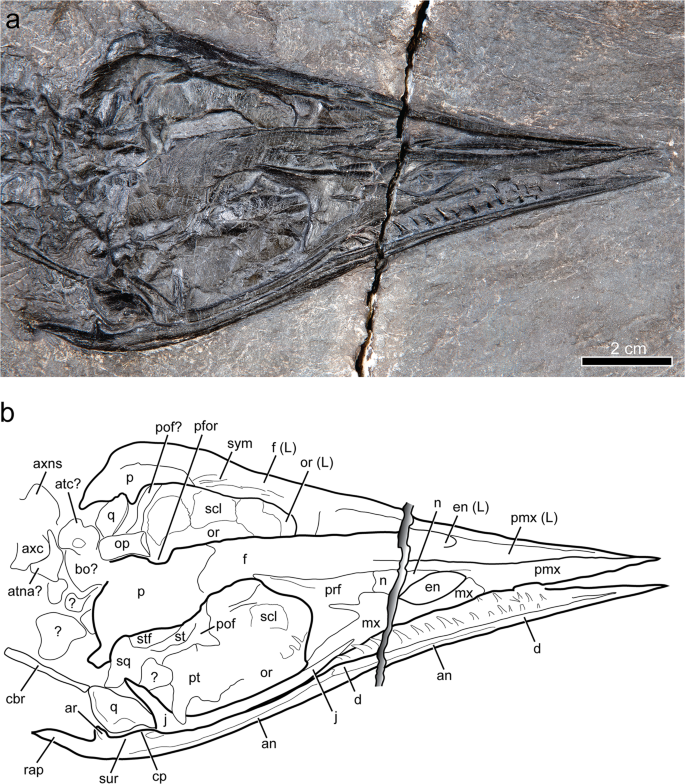
Skull of Gunakadeit joseeae. (a) Photograph. (b) Interpretation of the holotype specimen UAMES 23258 in oblique right lateral view. Abbreviations: an, angular; ar, articular; atc, atlas centrum; atna, atlas neural arch; axc, axis centrum; axns, axis neural spine; bo, basioccipital; cbr, ceratobranchial; cp, coronoid process; d, dentary; en, external naris; f, frontal; j, jugal; L, left; mx, maxilla; n, nasal; op, opisthotic; or, orbit; p, parietal; pfor, pineal foramen; pof, postorbitofrontal; pmx, premaxilla; prf, prefrontal; pt, pterygoid; q, quadrate; rap, retroarticular process; scl, scleral element; sq, squamosal; st, suptratemporal; stf, supratemporal fenestra; sur, surangular; sym, symphysis.

Jaw and teeth of Gunakadeit joseeae. (a) Photograph of right lower jaw. (b) Detail of single dentary tooth of the holotype specimen UAMES 23258 in right lateral view. Abbreviations: an, angular; cr, crown; d, dentary; er, enameled ridge; mx, maxilla.
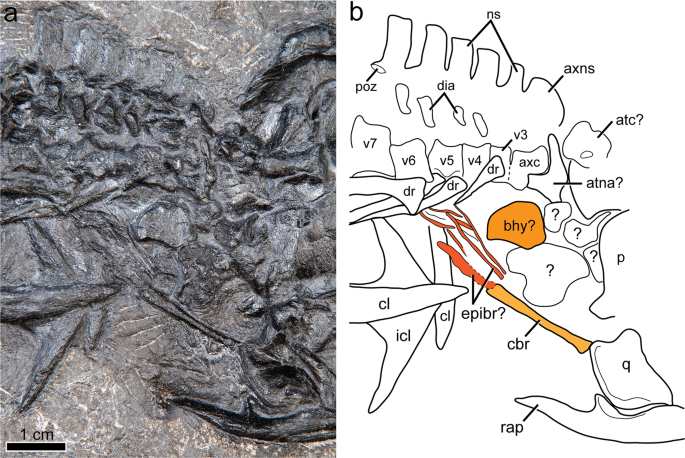
Cervical region and hyoid elements of Gunakadeit joseeae. (a) Photograph. (b) Interpretation of the holotype specimen UAMES 23258 in right lateral view. Colour scheme: light orange, ceratobranchial; dark orange, basihyal; reddish orange, epibranchials. Abbreviations: atc, atlas centrum; atna, atlas neural arch; axc, axis centrum; axns, axis neural spine; bhy, basihyoid; cbr, ceratobranchial; cl, clavicle; dia, diapophysis; dr, dorsal rib; epibr, epibranchial elements; icl, interclavicle; ns, neural spine; p, parietal; q, quadrate; rap, retroarticular process; v, vertebra.

Dorsal region and pectoral girdle of Gunakadeit joseeae. (a) Photograph. (b) Interpretation of the holotype specimen UAMES 23258 in right lateral view. Abbreviations: cl, clavicle; cor, coracoid; dc, dorsal centrum; dia, diapophysis; dr, dorsal rib; gast, gastralia; hum, humerus; int, intermedium; icl, interclavicle; mc, metacarpal; ph, phalanx; ns, neural spine; rad, radius; sc, scapula; ul, ulna; ule, ulnare.

Anterior caudal region and pelvic girdle of Gunakadeit joseeae. (a) Photograph. (b) Interpretation of the holotype specimen UAMES 23258 in right lateral view. Abbreviations: brom, bromalite; cc, caudal centrum; ch, chevron; fem, femur; fib, fibula; gast, gastralia; il, ilium; isch, ischium; L, left; mt, metatarsal; naf, neural arch facet; ns, neural spine; ph, phalanx; pub, pubis; R, right; sc, sacral centrum; sr, sacral rib; tar, tarsal; tib, tibia.
Type locality
Keku Islands of Southeast Alaska (Fig. 1). Precise coordinates are on file with the State of Alaska Office of History and Archaeology.
Age
Epigondolella postera Zone of the Hound Island Volcanics, middle Norian, Upper Triassic.
Measurements
(Supplementary Tables 1–3).
Diagnosis
Small-bodied (estimated total length 75–90 cm) thalattosaurian possessing the following autapomorphies: extremely pointed rostrum; posteriorly inclined dorsal process of the jugal; temporal region much shorter than orbit; anterior dentary edentulous; elongate retroarticular process; dorsal, sacral and anterior caudal neural spines with acute dorsal margin; elongate and narrow sacral ribs with distal expansion less than one third rib length; numerous, extremely fine gastralia; tapered posterodorsal process of ilium; tibia preaxial margin strongly convex. Gunakadeit joseeae can further be diagnosed on the basis of the following unique character combinations: large external naris; jugal extending far anterior of orbital margin; presence of supratemporal fenestra; reduced cervical region with approximately four cervicals; lateral processes of interclavicle more than 3 times longer than the width of the posterior process; proximal end of humerus wider than distal end; proximodistally elongated reniform radius; posterodistal process of the ilium short; tibia and fibula less than half femoral length.
Description and taxonomic comparisons
With the exception of non-fusion along the dorsal midline, the skull remains well articulated with a well-ossified scleral ring, although the visible portions of the braincase are largely disarticulated. Some neural arches have separated from the centra along the neurocentral suture. The carpus and to some extent the tarsus are poorly ossified, although this is conceivably related to their secondarily aquatic lifestyle. These features suggest that the holotype potentially represents a late juvenile or subadult, consistent with its small size (<1 m) compared to most other thalattosaurs.
The skull is triangular in dorsal view with converging lateral margins and lacks any ventral deflection of the rostrum (Figs. 2 and 3). The relative anteroposterior length of the external naris, measured as a percentage of preorbital length, is large in Gunakadeit (~20%) compared to typical askeptosauroids (e.g., 8% in Anshunsaurus huangguoshuensis9) (Supplementary Table 1). The temporal region is much shorter anteroposteriorly than the orbit; in all other thalattosaurians, the temporal region is nearly as long (e.g., Thalattosaurus alexandrae10) or longer (e.g., Askeptosaurus11) than the orbit.
The anterior ends of the premaxilla and dentary taper to an acuminate tip, resulting in a straight, narrow rostral morphology most similar to, but more pronounced than, askeptosauroids such as Askeptosaurus11 and Endennasaurus12. Xinpusaurus kohi13 also exhibits an acuminate rostrum, but differs in having a distinct overbite. It is not possible to tell if the anterior end of the premaxilla bears teeth; however, at least one premaxillary tooth is visible anterior to the external naris. The medial surface of the left premaxilla bears a longitudinal groove that extends for nearly its entire length anterior to the external naris, similar to Xinpusaurus xingyiensis14. The posteromedial process of the premaxilla forms the anterodorsal margin of the external naris and extends posteriorly to the frontals. The posteroventral margin of the premaxilla is overlapped by the maxilla at the anterior margin of the external naris. A shallow trough in the lateral surface of the premaxilla immediately anterior to the external naris results in a shelf that overhangs the anterodorsal margin of the external naris.
The maxilla bounds the entire ventral margin of the external naris, is anteroposteriorly longer than dorsoventrally tall and bears a posterodorsally ascending process overlapping the prefrontal, but not extending to the frontal, as in Thalattosaurus alexandrae10. A narrow posteroventral process lies along the entire ventral margin of the prefrontal and just contacts the anteroventral margin of the orbit. It is unclear if the maxilla bears teeth. The nasal separates the premaxilla from the maxilla along the posterodorsal margin of the external naris. Its dorsal margin is formed entirely by the premaxilla, thus excluding a contact between the nasals along the dorsal midline, a thalattosaurian synapomorphy15. Its anterior end tapers to the dorsal midpoint of the external naris differing markedly from T. alexandrae10, where the nasal forms the entire dorsal border of the external naris. The posterior end of the nasal abuts the prefrontal; it does not possess the narrow posterior process seen in most thalattosaurians. The prefrontal is robust and forms the entire anterior margin of the orbit. Ventrally, the prefrontal approaches, but does not contact the jugal, as in Clarazia16. A distinct, anteroposteriorly elongate ridge on its dorsolateral surface produces a prominent shelf of bone extending over the anterodorsal margin of the orbit. The lacrimal is absent. The gracile jugal bounds the entire ventral and posteroventral margins of the orbit. Its anterior end extends anterior to the orbit, similar to Clarazia16 but in contrast to Askeptosaurus11 and Miodentosaurus6 that terminate in line with, or posterior to the orbital margin, respectively. The dorsal process of the jugal is inclined posteriorly, not vertically, as in most thalattosaurians. The posterior process of the jugal is very reduced or possibly absent given that the dorsal process abuts the anterior margin of the quadrate similar to Xinpusaurus xingyiensis14 but in marked contrast to most askeptosauroids.
The poorly preserved postorbitofrontal appears to be a single ossification. It has a very short descending process to meet the very tall ascending process of the jugal, although it may be shortened due to breakage. The postorbitofrontal seemingly lacks an extensive anterior process, or it may be covered by the parietal. The anterior end of the frontal appears to be split by a posterodorsal process of the premaxilla, although poor preservation makes their exact relationships hard to discern. The frontal forms most of the dorsal rim of the orbit and bears a short, broad posterolateral process that may barely contact the postorbitofrontal. Along the dorsal midline, the medial surface of the frontal is visible and is surprisingly thick dorsoventrally (3–4 mm), bearing narrow, posteroventrally-sloping ridges along the symphysis, just anterior to the frontal-parietal suture. The frontal-parietal contact is sigmoidal. The parietal overlaps the frontal and encloses a prominent pineal foramen approximately in line with the postorbital bar. The anterolateral border of the parietal is formed by the supratemporal and postorbitofrontal and may contribute to the orbital border. A distinct posterolateral process is visible and appears to contact a distinct element, likely the squamosal, posterolaterally.
The anteroposteriorly elongated supratemporal is medial to, and overlaps the postorbitofrontal. The supratemporal extends anteriorly almost to the posterior margin of the frontal, possibly contacting it as in Clarazia17. The supratemporal forms the lateral border of a small but distinct opening interpreted to be the supratemporal fenestra, which is bordered medially by the parietal. The supratemporal fenestra is approximately 6 mm long and 2 mm wide, but not slit-like as interpreted for Thalattosaurus alexandrae10. Its position differs from askeptosauroids (e.g., Askeptosaurus) where it is bounded laterally by the postorbital and medially by the parietal, and other thalattosauroids (e.g., Thalattosaurus), where it is bounded laterally by the postorbitofrontal and medially by the frontal and supratemporal. Alternatively, the lateral border of the supratemporal fenestra may be formed entirely by the postorbitofrontal, which is difficult to distinguish from the supratemporal due to preservation. The quadrate abuts the posterior end of the jugal, exemplifying the extremely shortened temporal region. As preserved, the lateral surface is smooth, lacking the obvious ridges observed in Xinpusaurus suni18. The posterior margin is convex; however, much of its morphology is difficult to discern. Several small and disarticulated bones in the posterior skull region may be braincase elements. One of the larger elements near the atlas-axis complex may represent the basioccipital and another the left opisthotic. Exoccipital, prootic, supraoccipital and stapes were not identified.
A portion of the palate is visible through the right orbit. The lateral ramus of the pterygoid lies in proximity to the base of the dorsal process of the jugal. The exposed dorsal surface of the pterygoid is otherwise largely flat. The presence of palatal dentition (pterygoid or palatine) is unknown. Portions of both the left and right scleral rings are visible; the left ring includes at least three articulated, subrectangular scleral ossicles. Each ossicle thins toward the outer part of the ring and bears a radially striated surface texture. Based on ossicle proportions and the apparent diameter of the ring, there were likely fewer than the estimated fourteen scleral ossicles in Askeptosaurus11 or fifteen estimated in Concavispina19.
The left and right dentaries are fused along the midline symphysis for at least one third of their entire length, similar to Endennasaurus12 but unlike the shorter and unfused symphysis of Clarazia17. The anterior one-third of the dentary is clearly edentulous, an autapomorphy of this taxon. The dentary toothrow is straight. It contacts the surangular at a butt joint immediately posterior to the last visible tooth. The dentary-angular (or possibly the dentary-splenial) suture may occur approximately halfway along the length of the dentary. If this is the actual suture, the angular extends farther anteriorly than in most thalattosaurs, with the possible exception of Concavispina19. Posteriorly, the angular forms the ventral margin of the mandibular ramus at least as far as the mandibular fossa. The angular may also form some or all of the retroarticular process, although a distinct angular-articular suture is not visible. The retroarticular process is very elongate and dorsomedially deflected to a degree not seen in any other thalattosaur. The posterior margin of the mandibular fossa is marked by a mediolateral ridge; posterior to this the dorsal surface of the retroarticular process is relatively flat. The surangular forms the dorsal margin of the mandibular ramus and is bordered ventrally by the angular for its entire length. The coronoid process is a low mediolaterally thickened eminence, differing from the distinct and dorsally elongate process of Clarazia16. A distinct element identifiable as the coronoid is absent. A small, trough-like depression is located near the apex of the coronoid process. The posterior margin of the angular is marked by a suture visible just ventral to the glenoid fossa.
Fourteen teeth are visible in the right dentary and five in the left (Fig. 4). The dentition is homodont and each tooth consists of a slender, conical and recurved crown that is round in cross section. Tooth height increases posteriorly and the largest crowns are distributed throughout the posterior half of the toothrow, with the largest tooth being dentary tooth 13. Emergent replacement teeth are not visible. In the well-preserved eighth right dentary tooth, the crown comprises one-third of total tooth height and bears 4–5 straight enameled ridges from the base of the crow to the apex on the labial surface. A slight flattening on the apical surface of the crown may represent faint tooth wear. The surface of the root is smooth and expands in circumference basally. Roots are slightly mediolaterally flattened, presumably due to crushing of the pulp cavity. There is no evidence of distinct alveoli or a bony septum between adjoining teeth. Rather, the teeth appear to be set deeply against the medial surface of the jaw and thus tooth implantation is likely best described as pleurodont, as suggested for Clarazia16. It cannot be determined whether any type of shelf or lingual wall supports the roots, but some degree of bony attachment of teeth seems likely given that teeth show little sign of displacement.
A well-developed hyoid apparatus includes a prominent rod-like element, interpreted as ceratobranchial 1, and at least three additional thinner, rod-like elements preserved in close association just anterior to the clavicular arch (Fig. 5). Ceratobranchial 1 is minimally 18 mm in length (partially covered by the quadrate), straight, and slightly larger in circumference at its posterior end. Immediately posterior to it is another poorly ossified element, possibly epibranchial 1, which is 10 mm in length. Given its poor preservation it may have been only partly ossified, or even cartilaginous in life, as is typical of many modern amniotes20. At least two other poorly ossified and thin rod-like structures lie adjacent and parallel to epibranchial 1, possibly representing portions of neighboring hyoid arches. Lying in close proximity to these hyoid elements is an isolated plate-like bone that may represent the ossified corpus of the hyoid. Consistent with its identity is its overall shape, although it is not perfectly bilaterally symmetrical, and that it is apparently unpaired and lies in an appropriate relative position to other elements. In other thalattosaurians for which a hyoid apparatus is known (e.g., Anshunsaurus huangguoshuensis21; Concavispina19) only a single pair of elongate elements (i.e., certatobranchial 1) have been described.
The articulated axial column comprises 58 vertebrae, including a complete series of 32 presacral and three sacral vertebrae (Figs. 6 and 7). The incomplete tail preserves 23 partial vertebrae. Four cervical vertebrae, including axis and atlas, are identified; cervicals 3 and 4 are recognized on the basis of possessing short and straight ribs that are not long enough to have contacted the unossified sternum. The neck of Gunakadeit is very short compared to askeptosauroids such as Anshunsaurus wushaensis (15–16 cervicals22) or even short-necked thalattosauroids such as Hescheleria (7–8 cervicals23), but is comparable to Concavispina (~4 cervicals19). The atlas-axis complex is not fully articulated. The axis is identified by its anteroposteriorly expanded neural arch and attendant centrum. The atlas is represented by at least two elements lying immediately anterior to the axis and posterior to the skull; the larger element is possibly the atlantal centrum and a smaller, wedge-shaped atlantal neural arch. The identities of other smaller elements in this vicinity are uncertain. In lateral view, the axial centrum and its neural spine are anteroposteriorly longer than dorsoventrally tall. Atlantal and axial ribs were not identified, but the axis bears a prominent single-headed diapophysis on its ventrolateral surface.
The dorsal centra increase slightly in length in the anterior half of the dorsal series (from approximately 5 mm to 6–7 mm), and then remain relatively uniform in length into the sacrals. The ventral portion of the dorsal neural arches is robust and bears a single large, anteroventrally to posterodorsally elongate rib facet. The cervical and anterior dorsal neural spines are vertically oriented and evenly rectangular in outline, being approximately twice as tall as long, similar to Xinpusaurus suni24. However, throughout the posterior half of the dorsal series the neural spines become angled posteriorly and the apices of the neural spines are conspicuously pointed in lateral view, an autapomorphy of Gunakadeit. This contrasts with the dorsally-notched spines of Concavispina19, the short and broad spines of Askeptosaurus11 and the strongly constricted ventral portion of the neural spines in Anshunsaurus huangguoshuensis9. All of the visible rib heads are single-headed. The anterior-most rib associated with vertebra 3 is very short, being approximately two times the length of the centrum. On vertebra 4, the rib becomes longer (~20 mm) but remains relatively straight. The more posterior dorsal ribs are considerably longer (up to ~70 mm total length) and robust, are round in cross-section at midshaft, and distally flattened.
Three sacral vertebrae are distinguished by the presence of articulated sacral ribs on the first and third centra. Three sacrals are also found in Miodentosaurus25 and Concavispina19 while only two are present in Endennasaurus12 and Xinpusaurus sp.13. The sacral centra have similar proportions to the posteriormost dorsals, unlike Miodentosaurus where the sacrals are somewhat longer25. The sacral ribs are long, straight and possess a narrow, rounded shaft proximally. Distally, the ribs become dorsoventrally flattened and anteroposteriorly expanded. The width of the distal expansion is less than one third the entire length of the rib resulting in ribs that are relatively elongate, even compared to Endennasaurus12. The absence of a sacral rib on the second centrum may reflect primary absence or taphonomic loss.
Caudal centra are only slightly shorter than sacrals and posterior dorsals. A caudal rib may be associated with the first caudal centrum. The neural spines are narrowly rectangular in lateral view and display a slight anteroposterior expansion in the dorsal half of the spine. The spines are much taller than the height of the centrum, but not as comparatively tall as those in the Kössen Formation specimen (SMNS 90568) from Austria26. The lateral surfaces of the spines bear faint striations but lack the more prominent grooves seen in SMNS 9056826. The proximal spines are nearly vertically inclined but become posteriorly inclined and shorter, starting around caudals 6 and 7. The spine apex is not as pointed as those in the dorsal series. Chevrons on the first caudal vertebra are very long, more than two times the height of the centrum, but again not as comparatively long as those in SMNS 9056826, and diminish in length posteriorly.
The entire articulated gastral basket is preserved with the first gastralia appearing immediately posterior to the posterior end of the interclavicle and extending through the remainder of the torso, terminating just anterior to the pelvic girdle. Individual gastralia are extremely delicate and almost hair-like in appearance, making them difficult to precisely count. The gastralia are more gracile that those known from any other thalattosaurian, including the Kössen Formation specimen and Concavispina, which has approximately 100 pairs19. There appears to be a set of lateral elements, at least 70 in number, that overlap with a set of medial elements. Where visible, the lateral and medial elements are gently curved to nearly straight in shape. Two homogenous phosphatic masses are located adjacent to the pelvic girdle. Although lacking identifiable contents, their composition, shape and position suggest they are bromalites (gastric residues).
The specimen preserves both clavicles and coracoids, the interclavicle, and a partial right scapula (Fig. 6). The interclavicle is robust and T-shaped; however, the anteriormost tip is covered and it is unknown whether it possesses an anterior process, as in Clarazia17. The entire anterior margin is convex and the lateral processes are angled posteriorly at ~30 degrees. The lateral processes are conspicuously elongate; the length of each process is more than three times longer than the width of the mid portion of the posterior process, in contrast to most thalattosaurians (e.g., Endennasaurus12). The posterior process is mediolaterally widest at mid-length. The right clavicle is very long, being as long as the interclavicle and twice as long as the humerus, similar to Concavispina19, and is markedly robust midshaft, unlike Anshunsaurus huangguoshuensis21. The dorsal portion of the clavicle loosely contacts the scapula and terminates slightly beyond its posterior margin. As preserved, the scapula is anteroposteriorly longer than broad. The right coracoid is oval in shape, slightly anteroposteriorly longer than broad and is less than half as long as the interclavicle. The element is generally thin but appears slightly thickened along its posterolateral margin, in the vicinity of the glenoid. On its surface are a series of radiating, concentric lines. A coracoid foramen could not be identified.
The proximal end of the right humerus is wider than the distal end in contrast to askeptosauroids such as Miodentosaurus27. There appears to be no torsion in the shaft in contrast to Nectosaurus10, where the distal end of the humerus is twisted relative to the proximal end. A moderately developed deltopectoral crest may be present; however, it does not resemble the thin, flange-like crest seen in Thalattosaurus alexandrae10. Distally, the radial facet appears better developed than the ulnar facet and a shallow trough separates the two condyles. The well-developed radius and ulna are approximately 0.7 times the proximodistal length of the humerus. The radius is reniform, with a prominent convex preaxial margin, unlike the elongate radii seen in askeptosauroids. The similarly reniform radius of Xinpusaurus xingyiensis14 is proportionately shorter, being ~0.5 times the length of the humerus. The radius also lacks a distinct notch near its proximal end, as seen in Nectosaurus10. The proximal end of the ulna is much wider than the distal end, similar to Clarazia17 but it lacks a distinct olecranon process, in contrast to Nectosaurus10. The pre- and postaxial margins of the ulna are concave. Two proximal elements of the carpus are ossified; based on their position they may be the ulnare and intermedium. There appears to be a minimum of four digits present, but the rest of the autopodium is poorly preserved.
The ilium is curved in lateral view, with a well formed but short posterodorsal process that is less than two sacral vertebrae in combined length (Fig. 7). The dorsal margin is straight and lies in close proximity to the distal ends of the sacral ribs, but the ventral margin is gently curved, resulting in a tapering, wedge-shaped posterodorsal process, an autapomorphy of the taxon. In contrast, askeptosauroids such as Endennasaurus12 have nearly parallel-sided dorsal and ventral margins that only taper at the posteriormost end. In thalattosauroids such as Concavispina19 the posterior end of the posterodorsal process is slightly expanded in dorsoventral height and is also relatively longer, being more than twice the combined length of two sacrals. The acetabular end is the widest portion of the element. The acetabular surface is partly concealed but the lateral margin of the acetabulum is concave with a narrow, tapering anterior margin for the pubis and a ventrally deflected margin at the ischial end. What is tentatively identified as the pubis is situated ventral and anterior to the ilium and slightly overlaps the posteriormost gastralia. The acetabular end is robust and is set off from the thinner and anteroposteriorly expanded medial portion by a distinct shaft, which has a straight anterior margin and a concave posterior margin. An obturator foramen is absent. In contrast, the pubis of other thalattosaurians is typically mediolaterally shorter and more plate-like in overall appearance (e.g., Hescheleria17) and bears a distinct obturator foramen. Given the aberrant morphology of this element, it might alternatively represent a displaced sacral rib or another element. The ischium is largely not visible.
The femur is nearly identical in length to the humerus (Supplementary Tables 2 and 3) and paddle-shaped, with the distal end being conspicuously wider than the proximal end, similar to Thalattosaurus alexandrae10. The femur lacks any obvious proximal trochanters (internal or fourth), which are prominent in Miodentosaurus25 and Hescheleria17. The shaft is distinctly constricted and there is a midshaft depression that might be for insertion of the adductor musculature. The distal articular surface is evenly convex lacking distinct articular facets for the tibia or fibula. The tibia and fibula are about equal in length but less than half the length of the femur; in contrast, the tibia and fibula are approximately half as long as the femur in Clarazia17 or more than half as long in Anshunsaurus huangguoshuensis21. The tibia possesses an autapomorphic convex preaxial margin. The fibula is broadly expanded midshaft but both pre- and postaxial margins are convex. Its distal end is only slightly more expanded than its proximal end; in contrast, the distal end of the fibula is markedly expanded in Clarazia17. Six or seven elements are visible in the tarsus, but are poorly preserved and/or ossified. The largest is the astragalus, which lies distal to the spatium osseum and in contact with both the tibia and fibula. A small unidentified element is located preaxially of the astragalus; distal to these proximal tarsals are three (possibly four) distal tarsals. A small ossicle located posterodistal to the astragalus may be the calcaneum. Only two probable metatarsals (I and II?) are present and there are 12 proximal phalanges still in articulation, indicating five digits in the hind limb.
Phylogenetic position
After the deletion of three highly incomplete taxa (see Methods) our strict consensus tree (Fig. 8) is nearly completely resolved, with the exception of a polytomy among Anshunsaurus huanggoushuensis, A. wushaensis and Miodentosaurus. A monophyletic Thalattosauria is recovered with a basal dichotomy splitting all ingroup OTUs into one of two traditionally recognized clades, Askeptosauroidea and Thalattosauroidea, in accordance with most previous analyses. Gunakadeit joseeae is recovered as the basalmost member of Thalattosauroidea. Bootstrap values indicate strong support for the monophyly of Thalattosauria (99%) and Thalattosauroidea (88%); however, support for internal branches is generally less than 50%, with the exception of the sister group relationship between Thalattosaurus alexandrae and T. borealis and Xinpusaurus kohi and X. suni (both 58%).
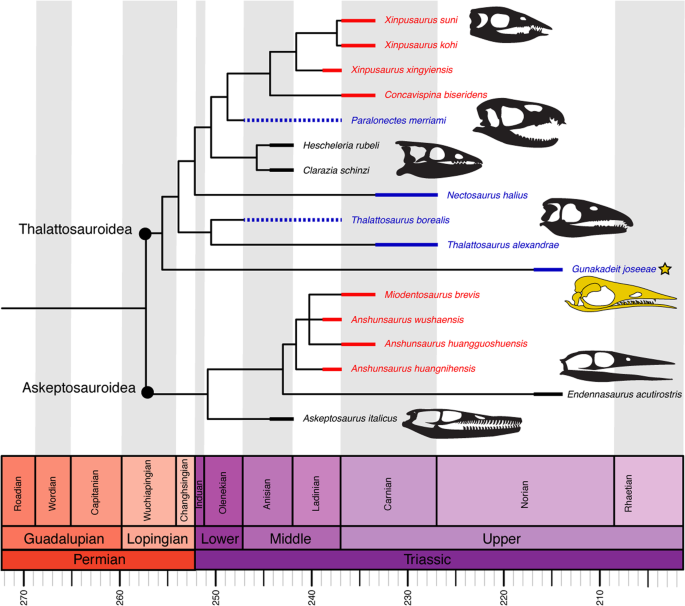
Time-scaled strict consensus topology of Thalattosauria. Thickened lines represent age range of each taxon; dashed line represents range of uncertainty. Colour scheme: red, Eastern Tethys (modern China); blue, Western Tethys (modern Europe); black, eastern Panthalassa (modern western North America).
Source: Ecology - nature.com
