Growth stages of development are the longest and most variable
To test the prediction that growth periods represent the longest and most variable phases of offspring development, we conducted a fine-scale analysis of developmental rates in a taxonomically diverse set of species (n = 20) with existing information on the timing of key developmental stages (Supplementary Fig. 1 and Supplementary Table 1). Specifically, we examined four distinct phases in avian ontogeny spanning both pre-hatching (incubation) and post-hatching (fledging) periods (Fig. 1a). Phases 1 and 2, defined on the basis of Hamburger–Hamilton (HH) stages 1–24 and 25–32, respectively, correspond to periods of chick embryogenesis. These early stages of prenatal development consist primarily of cell differentiation and embryo formation rather than absolute growth28, and we therefore consider these as “non-growth” phases. In contrast, phases 3 and 4, corresponding to HH33 to hatching and hatching through to fledging, are primarily concerned with periods of prenatal (phase 3) and postnatal (phase 4) growth of existing structures (see Supplementary Methods for extended justification of these developmental phases). We used this framework to investigate variation in the duration and partitioning of avian offspring development.
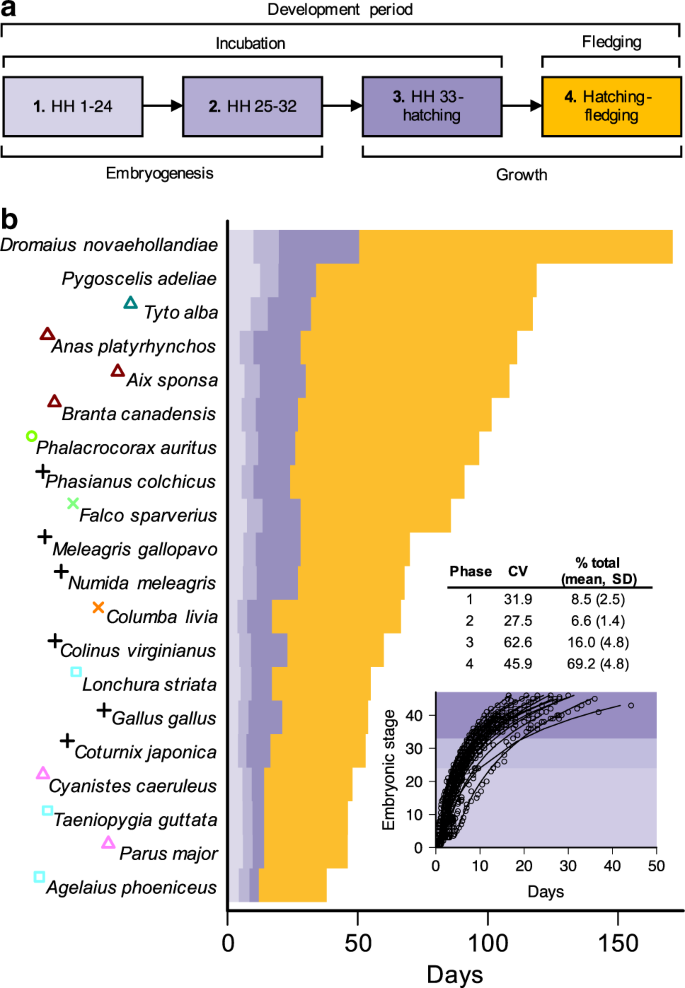
a Schematic illustrating four distinct phases of avian ontogeny. Phase 1 and phase 2 corresponding to Hamburger–Hamilton stages (HH) 1–24 and 25–32, respectively, represent embryonic developmental stages primarily associated with embryogenesis (i.e., non-growth). In contrast, phases 3 (HH33 to hatching) and 4 (post-hatching fledging period) correspond to developmental periods consisting largely of growth. b Stacked bar chart showing time intervals associated with phases 1 to 4 for 20 bird species with available information on the timing of embryonic developmental stages, with species are ordered by total developmental duration. Coloured symbols next to species names correspond to the major taxonomic groups identified in Fig. 2. Inset graph shows the staging data and fitted curves used to estimate the time points separating phases 1–3. Inset table reports the coefficient of variation (CV) and percentage of total developmental period length (% total) accounted for by each of the four phases. Source data are provided as a Source Data file.
We found that the durations of developmental stages associated with embryogenesis (phases 1 and 2) account for only a small proportion of the variance in overall developmental duration across species (Fig. 1b). At these early stages of development, all bird embryos—regardless of species identity and eventual adult body size—are of comparatively similar size and therefore expected to have approximately similar growth rates7. In contrast, the durations of growth phases (phases 3 and 4) are longer and more variable than non-growth phases and account for a far greater proportion of the variance in developmental duration among species (Fig. 1b). The longer duration of growth phases relative to embryogenesis phases is consistent with the well-characterised phenomenon of declining growth rates over ontogeny, caused by decreases in the ratio of energy acquisition to energy loss as developing organisms increase in size5. Furthermore, greater variance in the duration of growth phases relative to non-growth phases (as indicated by coefficient of variation scores; Fig. 1b) is also consistent with greater size-related effects on the later stages of development. As development progresses, offspring of different species become increasingly different in size and therefore exhibit far greater disparity in relative growth rates compared with earlier stages of development.
Developmental durations are phylogenetically conserved
Our observation that the developmental phases associated primarily with growth are longer and more variable than earlier non-growth phases predicts that body size should explain a significant amount of variation in developmental durations due to metabolic scaling rules. To test this idea more broadly, we collected data on developmental durations (incubation and fledging periods) for 3096 bird species covering the breadth of the avian phylogeny (Fig. 2a and Supplementary Fig. 2). In our dataset, overall developmental durations ranged from ~20 days in some passerine species (e.g., Volatinia jacarina) to >350 days in some seabird lineages (e.g., Diomedea). Likewise, estimates of the proportion of development allocated to incubation relative to fledging (i.e., the incubation fraction) also varied markedly across species, ranging from ~0.15 (e.g., Struthio camelus) to >0.95 in certain landfowl and shorebird species (e.g., Megapodius pritchardii and Synthliboramphus wumizusume).
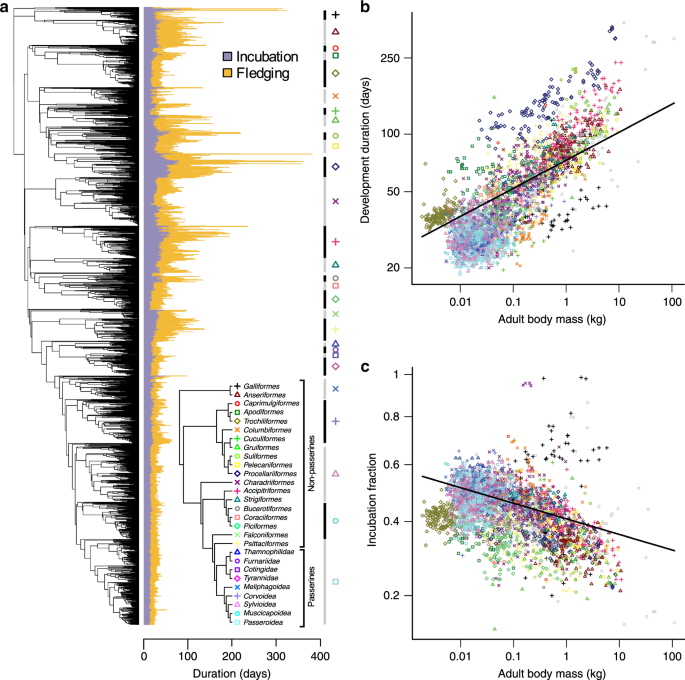
a The phylogenetic distribution of incubation, fledging and total development (incubation + fledging) period across 3096 species of birds. Inset tree schematic indicates the relationships among major taxonomic groups (>20 spp.) and provides a key for the plotting symbols used throughout the figure. b, c Allometric relationships of (log-transformed) development period length (b) and (square root-transformed) incubation fraction (c) with (log-transformed) adult body mass. Lines indicate the regression slopes estimated by phylogenetic regression. Source data are provided as a Source Data file.
Before addressing relationships with body size, we first quantified the extent of phylogenetic signal in avian developmental durations. Fitting Pagel’s λ model29 to our dataset, we found that developmental variables exhibited strong phylogenetic signal, with λ values [95% confidence interval (95% CI)] of 0.93 [0.91, 0.94] and 0.86 [0.83, 0.89] for developmental duration and incubation fraction, respectively. This reflects a pervasive pattern in our dataset, that species within clades tend to exhibit similar developmental durations and incubation fractions (Fig. 2a), such that on average closely related species have more similar trait values than more distantly related species.
Body size explains less variation than previously thought
Against this backdrop of evolutionary conservatism, we used phylogenetic regression30 and variance partitioning techniques31 to test the relationships between body size and avian developmental duration while jointly estimating phylogenetic effects, and to compare the contributions of predictor variables (body size) and variance components (phylogenetic effects) to the overall fit of the model. Using this approach, as predicted we found that overall developmental duration is positively related to body size across bird species (Fig. 2b). Furthermore, for incubation fraction we found that the offspring of larger-bodied species have proportionally shorter incubation periods relative to fledging periods (Fig. 2c), presumably reflecting energetic and/or ecological constraints associated with laying and/or developing in larger eggs. In both cases, we used adult body mass values as our index of body size across species, which we consider to represent a useful albeit imperfect proxy for offspring size at the end of development. However, we note that results were similar when we use an alternative proxy for offspring size (initial egg mass) (Supplementary Table 2). Variance partitioning revealed that the partial R2 values associated with these phylogenetically-adjusted allometric relationships were 0.22 and 0.05 for developmental duration and incubation fraction, respectively. In contrast, the partial R2 values associated with the phylogenetic (covariance) components of each model were far greater: 0.79 and 0.59, respectively.
The significant relationships we observe between body size and developmental durations are in line with our predictions based on embryological data and provide broad empirical support for the role of size-related constraints in determining both the duration and partitioning of avian developmental periods6,7,8. However, after accounting for phylogenetic effects, we found that the importance of body size for explaining variation in developmental durations across birds was surprisingly low, particularly considering that early tests implied that as much as 85% of interspecific variation in incubation period could be explained by body size effects32,33,34. In contrast, our comparatively low estimates for the variance explained by body size (5–22%) support the conclusion that, although important, allometric constraints play a more minor role in determining the length and partitioning of avian developmental periods than once thought21. Instead, our quantitative estimates indicate that a greater proportion of the variance in avian developmental durations is attributable to phylogenetic history rather than body size. This finding is apparent in the observation that species within clades typically share similar developmental duration values that are largely unrelated to variation in body size both within and between clades (Fig. 2 and Supplementary Fig. 3).
Ecology predicts broad-scale variation in development periods
The existence of substantial mass-independent differences in developmental periods among bird lineages is intriguing, as it raises questions regarding the relative importance of mechanistic versus ecological constraints in generating interspecific diversity in avian developmental periods. However, such questions have yet to be addressed at broad scales21. The idea that ecology plays an important role in driving the evolution of developmental periods is rooted in classical life-history optimisation theory2,4,9. To test the relative importance of ecology in explaining broad-scale variation in developmental periods, we studied the individual and combined effects of 16 variables related to behavioural, ecological, environmental, and life-history variation across species (plus body size) that have previously been linked to patterns of selection acting on avian developmental periods10,11,12,13,14,15,16,17,18,19,20 (Fig. 3). Specifically, we used phylogenetic generalised least squares (PGLS) regression with optimised Pagel’s lambda29 to test for relationships between ecological traits and developmental duration that are predicted by adaptive hypotheses. This approach, which is based on an underlying Brownian motion (BM) model, is suitable for testing for evolutionary associations between variables across species while controlling for the degree of phylogenetic non-independence in the data35. As such, this represents an appropriate framework within with to address our hypotheses as we are able to robustly test for correlations between ecological traits and developmental durations that are predicted by adaptive hypotheses, while simultaneously estimating and correcting for the degree of phylogenetic effects31,35. Furthermore, we performed a model comparison analysis and found that the lambda model greatly outperformed alternative candidate models [strict BM, Ornstein-Uhlenbeck (OU)] for the phylogenetic covariance structure of the residuals of our models and therefore represents the most appropriate statistical model with which to analyse our dataset.

Phylogenetically controlled multi-predictor models of development period, incubation fraction, incubation period and fledging period. Unfilled circles indicate factors that were significant as single predictors but not significant in a multi-predictor model. Gaps indicate factors that were not significant (ΔAIC < 2) as single predictors and were therefore not included in the multi-predictor model. Red and blue points indicate predictors with positive and negative effects, respectively. Predictors with grey points (e.g., Diet) represent categorical variables with >2 (‘multi’) levels. ΔAIC values indicate the change in model support when the focal predictor was dropped from the model, with larger ΔAIC values indicating greater support for the importance of a predictor. Sample sizes (number of species) for the models were 1665, 1685, 1935 and 1665 for development period, incubation fraction, incubation period and fledging period, respectively.
Our analyses revealed several important correlates of variation in avian developmental durations. First, after testing each predictor separately (see Supplementary Figs. 4–7), we found strong relationships between several variables and developmental duration and incubation fraction across species (Supplementary Table 2). By combining all significant single predictors in multi-predictor models, we were then able to identify sets of important predictors with unique effects that are independent of phylogeny. We found that, in addition to being larger, species with longer overall developmental durations tend to be longer lived, with smaller clutches, biparental care, elevated nest heights, vertebrate-eating/scavenging dietary niches, and pelagic foraging ecologies (Fig. 3 and Supplementary Table 3). These species also tend to be non-migratory and have more equatorial and insular breeding-range distributions. For incubation fraction, in addition to the negative relationship with body size, we found that species with proportionally longer incubation periods tend to have uniparental parental care, are typically insectivorous and nocturnal, and have more polar breeding-range distributions (Fig. 3 and Supplementary Table 4). In both cases, broadly similar effects were found using initial egg mass as an alternative proxy for body size (Supplementary Fig. 8 and Supplementary Tables 5 and 6). Partial R2 values for these models indicated that, after controlling for phylogenetic and body size effects, the unique effects of ‘ecological’ variables included in multi-predictor models accounted for ~12% and ~4% of the variance in developmental duration and incubation fraction, respectively (Table 1). Interestingly, the magnitude of these effects were similar to those associated with body size (Table 1), implying that ecological and allometric effects (as measured here) explain roughly equivalent proportions of variation in developmental durations among bird species. Nonetheless, the variance associated with phylogenetic components indicated that phylogenetic effects remained a dominant source of variation in these models (Table 1). In total, these models incorporating body size, ecological, and phylogenetic effects accounted for 62–93% of the variation in developmental durations across species.
These results have several important implications. Most notably, they show that behavioural and ecological variables among species are significant predictors of variation in developmental durations across species, consistent with an important role for ecology in driving the evolution of avian developmental durations4. In particular, three main ‘ecological syndromes’ appear to be associated with variation in developmental durations. First, longer developmental durations are generally associated with factors that presumably increase the safety of the developmental environment from predation threat or other mortality risks, such as nesting in relatively inaccessible sites (nest height), on islands (insularity), or having more than one parent to provide for and protect the offspring (biparental care). The idea that nesting in safe places may relax selection for rapid development is consistent with work by Remeš and Martin14, who found nestling growth rates to be positively associated with predation rates across passerines. Second, factors linked to phenological effects, such as breeding at temperate latitudes, insectivory, and migratory ecology, tend to be associated with shorter developmental durations. In species where reproductive success is driven largely by an individual’s ability to coincide their reproduction with peak seasonal food availability4,36,37, the need to operate within a tight timeframe to avoid phenological mismatch is likely to select for rapid development38. Third, several of the patterns we observe are also consistent with the importance of trade-offs between reproduction and survival for determining variation in avian developmental strategies. For example, shorter developmental periods in species with short lifespans and large clutches are consistent with selection for ‘fast’ life-histories and greater investment in reproduction (independent of body size)39,40, whereas longer developmental periods among species with vertebrate hunting/scavenging diets are potentially explained by selection for slower development to mitigate costs associated with limited and/or unpredictable food availability41,42.
Furthermore, by considering predictors of incubation and fledging period separately, our results provide further insight into the patterns of selection generating underlying divergence in overall developmental duration and pre- versus postnatal allocation (Fig. 3 and Supplementary Tables 7–10). For instance, our finding that nocturnal species have larger incubation fractions than diurnal species is seemingly driven by nocturnal species having relatively long incubation periods rather than particularly short fledging stages. This makes sense if lower daytime parental activity disproportionally reduces nest predation risk during the incubation period relative to the fledging period19, thus relaxing selection for rapid development inside the egg. Similarly, the longer developmental periods of pelagic species are largely driven by relatively long incubation periods, which may be a consequence of selection for advanced development at hatching15 or lower rates of egg predation due to inaccessible breeding locations40.
In contrast, our results show that species with uniparental care tend to have overall shorter developmental durations (and greater incubation fractions) largely because of reduced fledging durations. This is consistent with predictions for evolutionary associations between single parent care and short post-hatching offspring development periods4,43, but the direction of causality remains unclear. On the one hand, uniparental care may generate selection for rapid post-hatching offspring development to reduce the burden of care, but on the other hand short post-hatching periods may facilitate desertion by one of the parents (typically the male), implying a reversal in the direction of cause and effect44.
Finally, our results challenge several assumptions regarding relationships between developmental durations and other factors at broad scales. In particular, ambient climate is predicted to shape broad-scale patterns of developmental rates in birds via its effect on egg temperature and parental behaviour21. However, after controlling for the effect of other factors, we found no evidence that variation in environmental conditions (temperature and precipitation) was related to developmental duration across species. This finding supports the view that offspring are to a large extent buffered from variation in ambient environmental conditions by parental adaptations such as nest design, incubation efficiency and provisioning rate17,45,46.
Surprisingly, we also found no significant relationships with developmental mode (precocial, semi-precocial, altricial) or nest type (cavity, closed, open, mixed). This is despite strong expectations for significant associations26,47,48,49 and seemingly large differences between groups in the raw data (see Supplementary Figs. 4–7). We attribute these negative results to the effect of correcting for phylogenetic non-independence among species in our models. Variation in developmental mode and nest type are phylogenetically conserved across the avian phylogeny50. Power to detect significant relationships with traits that have independently evolved only a few times is limited and so their effects cannot be disentangled from underlying patterns of shared evolutionary history and/or ecology31. Greater clarity on whether factors such as developmental mode and nesting behaviour directly influence the evolution of developmental durations or are simply associated at broad scales via phylogenetically conserved constraints will likely come from integrating data on equivalent traits from other groups (e.g., all vertebrates) to generate sufficient independent phylogenetic replication to conclusively test these relationships.
Conclusions
Overall, our study reveals key drivers of developmental durations across the breadth of the avian phylogeny, providing broad, quantitative insight into the relative importance of mechanistic constraints and ecologically mediated selection in explaining variation in key life-history traits. Furthermore, our results highlight the pervasive impact of phylogenetic history in shaping variation in species’ developmental durations. The close association between developmental duration, species’ traits and phylogeny implies a strong signal of evolutionary conservatism, both in terms of species’ developmental durations and the combinations of factors (‘syndromes’) that co-evolve with them, echoing the conclusions from other large-scale phylogenetic analyses of avian life-history traits40,50. Although birds provide sufficient evolutionary replication to investigate the importance of many factors, phylogenetic constraints and evolutionary conservatism makes it difficult to tease apart the effects of other, less labile, traits. Thus, a potentially fruitful avenue of future research would be to address these questions over even broader phylogenetic scales to better address the effects of body size, species’ traits and phylogenetic constraints (e.g., mutation rates) on the evolutionary diversification of developmental durations.
Source: Ecology - nature.com
