Family Unionidae Rafinesque, 1820
Subfamily Unioninae Rafinesque, 1820
Tribe Anodontini Rafinesque, 1820
Genus Anodonta Lamarck, 1799
=Colletopterum Bourguignat, 1880 (type species: Anodonta letourneuxi Bourguignat, 1870 = Anodonta anatina; subsequent designation by Simpson, 1900)25.
=Piscinaliana Bourguignat, 1881 (type species: Anodonta piscinalis Nilsson, 1823 = Anodonta anatina; subsequent designation by Locard (1890); unavailable name, primary junior homonym of Piscinaliana Paladilhe, 1866, Gastropoda, Valvatidae)25.
Type species: Mytilus cygneus Linnaeus, 1758 (monotypy).
Diagnosis. Anodonta is very similar to Pseudanodonta but can be distinguished by a broadly rounded anterior margin, a shorter hinge length from the umbo to the ligament’s posterior limit45, and shorter and less closely spaced papillae of the inhalant siphon46.
Distribution. Two species of this genus are recorded from Russia (Fig. 2A–D and Table 1). Anodonta anatina inhabits European Russia, Urals, and Siberia eastwards to the Lena River basin (Fig. 6D), while the range of A. cygnea is restricted to water bodies of the Baltic, Black, Azov, and Caspian Sea drainage basins (Fig. 6E).
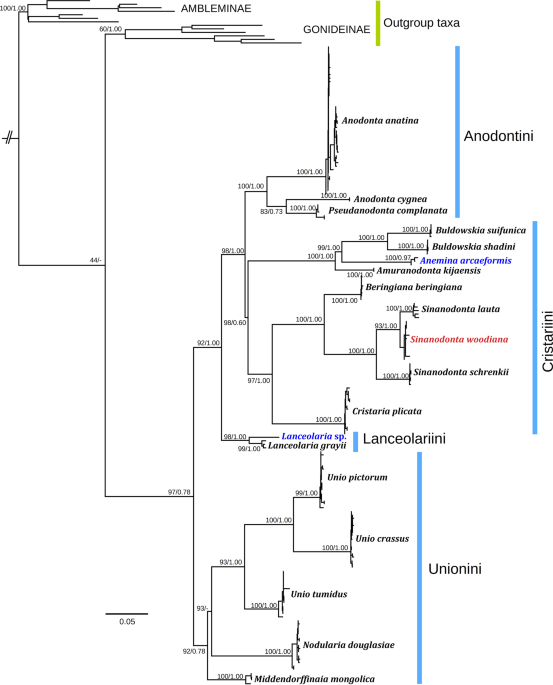
Comments. Phylogenetically, this genus-level clade appears to be paraphyletic because it contains Pseudanodonta complanata lineage (Fig. 1). The taxonomy of these two genera has not been discussed due to insufficient molecular data on Anodonta species from North America.
Genus Pseudanodonta Bourguignat, 1876
Type species: Anodonta complanata Rossmässler, 1835 (subsequent designation by Westerlund, 1902)25.
Diagnosis. See above.
Distribution. Pseudanodonta complanata (Fig. 2E) inhabits rivers of the Baltic, Black, Azov, and Caspian Sea drainage basins (Table 1 and Fig. 6E).
Comments. This genus may represent a junior synonym of Anodonta.
Tribe Cristariini Lopes-Lima et al . , 2017
Genus Amuranodonta Moskvicheva, 1973
=Amurbuldowskia Bogatov & Starobogatov, 1996 (type species: Buldowskia (Amuranodonta) boloniensis Zatravkin & Bogatov, 1987; original designation)22,25.
Type species: Amuranodonta kijaensis Moskvicheva, 1973 (original designation).
Diagnosis. This genus is most similar to Buldowskia, Anemina s. str., and Beringiana but can easily be distinguished from these taxa by an elongated shell with a characteristic elongated, triangular posterior margin (vs ovate or rounded shell with rather rounded posterior margin).
Distribution. This genus (Fig. 3G,H) seems to represent a lineage endemic to the Amur Basin (Fig. 6C and Table 1). It is also known to occur in the Chinese part of the Ussuri River47.
Comments. This monotypic genus represents a lineage that is phylogenetically distant from the Buldowskia and Anemina s. str. clades (mean COI p-distance = 15.0% and 14.6%, respectively) (Fig. 1). Our first reviser action on the precedence of simultaneous synonyms: Amuranodonta kijaensis over A. parva (see Supplementary Note for explanation).
Genus Beringiana Starobogatov in Zatravkin, 1983
=Kunashiria Starobogatov in Zatravkin, 1983 (type species: Anodonta japonica Clessin, 1874; original designation)25,48.
=Arsenievinaia Zatravkin & Bogatov, 1987 (type species: Amuranodonta sihotealinica Zatravkin & Starobogatov, 1984; original designation)20,25.
Type species: Anodonta cellensis var. beringiana Middendorff, 1851 (original designation).
Diagnosis. This genus can be distinguished from Amuranodonta by a less elongated, inequilateral shell with a rounded posterior margin and from Buldowskia by a comparatively weakly pronounced umbo. Both Cristaria and Sinanodonta have much higher, elliptical shells, although several morphological varieties of B. beringiana from small lakes have short, rounded shells with broad posterior margin (Fig. 4H).
Distribution. Beringiana beringiana occurs in Russia, being widespread in rivers and lakes east of the Lena Basin, coastal rivers of the Japan and Okhotsk Sea drainage basins northeast of Vladivostok (from the Kievka River), and on the North Pacific Islands (Kuriles and Sakhalin) (Fig. 6B and Table 1). This is a conchologically variable species, with the largest number of synonyms introduced for its morphotypes from different water bodies of the Russian Far East (Fig. 4F–H and Supplementary Note). Outside Russia, this species is known from Alaska, Western Pacific Region, and Canada in North America, and from Japan44,49.
Comments. Williams et al.49 assumed that Beringiana is a junior synonym of Sinanodonta, but this genus represents a distant genus-level phylogenetic lineage and the two are here treated as distinct taxa (Fig. 1). Kunashiria is a synonym of Beringiana, because its type species, B. japonica, belongs to the latter genus44.
Genus Buldowskia Moskvicheva, 1973
Type species: Anodonta arcaeformis var. suifunica Lindholm, 1925 (original designation).
Diagnosis. This genus can be distinguished from Amuranodonta by an ovate shell with a rounded posterior margin, and from Beringiana by a comparatively pronounced umbo, while in some lacustrine populations of Buldowskia shadini the umbo is not pronounced (Fig. 3E). Buldowskia and Anemina s. str. are almost indistinguishable morphologically.
Distribution. Two Buldowskia species are recorded in Russia (Fig. 3A–F and Table 1). This genus is distributed in the Amur and Razdolnaya River basins, and in the coastal rivers southwest of Vladivostok (Fig. 6A,C).
Comments. This genus is phylogenetically distant from the Amuranodonta and Anemina s. str. clades (mean COI p-distance = 15.0% and 16.7%, respectively) (Fig. 1). In addition to Buldowskia suifunica and B. shadini, it contains two lineages endemic to South Korea, i.e. B. flavotincta (Martens, 1905) and B. iwakawai (Suzuki, 1939), and a species new to science from Japan44,50. Our first reviser action on the precedence of simultaneous synonyms: Buldowskia shadini over B. fuscoviridis and B. buldowskii (see Supplementary Note for explanation).
Genus Cristaria Schumacher, 1817
Type species: Cristaria tuberculata Schumacher, 1817 (monotypy).
Diagnosis. Cristaria species can be distinguished from those of Sinanodonta by the presence of reduced lateral teeth, which Sinanodonta species lack, the absence of pseudocardinal teeth, and a comparatively thick shell. Additionally, Cristaria species have a well-developed dorso-posterior keel and a more angulate, higher shell with clear angle between dorsal and posterior margin, although Sinanodonta schrenkii often has an angulate shell with clear angle between dorsal and posterior margin.
Distribution. Cristaria plicata was recorded throughout the Amur Basin, and in the Tym’ River, central Sakhalin51 (Figs. 4E, 6C and Table 1). There are several records of subfossil shells of this species from the Pleistocene deposits in the Tym’ Valley6,52.
Genus Sinanodonta Modell, 1945
=Cristariopsis Moskvicheva, 1973 (type species: Sinanodonta (Cristariopsis) crassitesta Moskvicheva, 1973; original designation)19,25.
=Ellipsanodon Bogatov & Starobogatov, 1996 (type species: Sinanodonta (Ellipsanodon) ovata Bogatov & Starobogatov, 1996; original designation)21,25.
Type species: Symphynota magnifica Lea, 1834 (by typification of a replaced name)25.
Diagnosis. This genus can be distinguished from Cristaria by the lack of lateral teeth and a comparatively thin, rather fragile shell. Usually, Sinanodonta taxa have more ovate shells with a rather rounded angle between dorsal and posterior margin and a weakly developed or lacking dorso-posterior keel. However, Sinanodonta schrenkii often has an angulate shell with clear angle between dorsal and posterior margin.
Distribution. Two native Sinanodonta species are recorded in Russia (Fig. 2F–H and Table 1). This genus is distributed in the Amur and Razdolnaya basins, and in the coastal rivers southwest of Vladivostok (Fig. 6A,B). The non-native species Sinanodonta woodiana was recorded from the Yenisei River, in which it was found in a thermally polluted river channel in sympatry with an introduced population of S. lauta10 (Fig. 6A).
Tribe Lanceolariini Froufe et al . , 2017
Genus Lanceolaria Conrad, 1853
=Cylindrica Simpson, 1900 (type species: Nodularia cylindrica Simpson, 1900; original designation; unavailable name, primary homonym of Cylindrica Clessin, 1882, Gastropoda, Hydrobiidae)25.
=Pericylindrica Tomlin, 1930 (replacement name for Cylindrica Simpson, 1900)25.
=Prolancealaria Moskvicheva, 1973 (type species: Unio grayii Griffith & Pidgeon, 1833; original designation)7,25.
Type species: Unio grayanus Lea, 1834 (monotypy).
Diagnosis. This genus can easily be distinguished from the other Russian Unionidae by its unique lanceolate shell shape.
Distribution. Lanceolaria grayii inhabits Lake Khanka, Ussuri Basin, and Lower Amur (Figs. 4D, 6A and Table 1), representing the most northern enclave for this remarkable lineage of the Yangtze Basin fauna44,53.
Tribe Unionini Rafinesque, 1820
Genus Middendorffinaia Moskvicheva & Starobogatov, 1973
=Suifununio Moskvicheva & Starobogatov, 1973 (type species: Middendorffinaia (Suifununio) suifunensis Moskvicheva & Starobogatov, 1973; original designation)18,25.
=Pseudopotomida Moskvicheva & Starobogatov, 1973 (type species: Middendorffinaia (Pseudopotomida) shadini Moskvicheva & Starobogatov, 1973; original designation)18,25.
Type species: Unio mongolicus Middendorff, 1851 (original designation).
Diagnosis. This monotypic genus is externally similar to Nodularia, from which it can be distinguished by a higher, shorter shell, a strongly convex or even angulate hinge plate, more convex dorsal margin, and fine umbonal sculpture with small regular tubercles and narrow ridges in young shells. However, umbonal sculpture is often weakly developed or absent.
Distribution. Middendorffinaia mongolica is distributed in the Amur and Razdolnaya river basins, and in coastal rivers west of Nakhodka54 and southwest of Vladivostok (Figs. 5A–D, 6G and Table 1). The record from a coastal river of the Okhotsk Sea drainage basin40,55 is Nodularia douglasiae (Fig. 5F).
Comments. Middendorff (p. 277)12 described his Unio mongolicus based on a single specimen, which must be considered the holotype (by monotypy). The type locality was stated as follows: “Aus einem Gebirgsbache ohnfern Gorbitza in Daurien” [Russia, Transbaikalia, a mountain spring near Gorbitsa village (53.1027°N, 119.2169°E)]. The holotype was lost a long time ago, at least before 197318,20,24,25. The two figures of Middendorff12 show outside of the left valve and dorsal side of the shell. A relatively high shell with convex dorsal margin and umbo situated near the anterior margin indicates that Unio mongolicus sensu Middendorff is a distinct species, not a synonym of Nodularia douglasiae.
Zhadin5 assumed that Unio mongolicus sensu Middendorff is a rare member of the Margaritiferidae, and placed it in the genus Margaritana Schumacher, 1817. However, in his later work this species was called Unio douglasiae var. mongolicus with a question mark6. Moskvicheva and Starobogatov18 identified several specimens from the Ussuri Basin as prospective representatives of Unio mongolicus, but their specimens belong to Nodularia douglasiae (Fig. 5G). A new genus, Middendorffinaia, was established, with Unio mongolicus Middendorff as its type species18. This genus included three subgenera: Middendorffinaia s. str., Pseudopotomida, and Suifununio. Taxa placed within Middendorffinaia s. str., with exception of Unio mongolicus sensu Middendorff, belong to Nodularia douglasiae. In contrast, the Pseudopotomida and Suifununio species represent conchological varieties of Unio mongolicus sensu Middendorff (Supplementary Note). Two more such varieties are described as separate nominal species7,24.
Graf26 placed Middendorffinaia s. str. taxa as synonyms of Unio crassus mongolicus Middendorff, and Pseudopotomida and Suifununio taxa as synonyms of Inversidens pantoensis (Neumayr, 1899). This point of view highlighted differences between Unio mongolicus sensu Middendorff (with Pseudopotomida and Suifununio taxa) and U. mongolicus sensu Moskvicheva and Starobogatov (with their additional Middendorffinaia s. str. taxa belonging to Nodularia douglasiae). However, Unio mongolicus Middendorff with its varieties (Pseudopotomida and Suifununio spp.) is phylogenetically and morphologically distant from both the European Unio and East Asian Inversidens56,57.
Klishko et al.40 followed the concept of Unio mongolicus sensu Moskvicheva and Starobogatov18 and pictured a Nodularia douglasiae shell collected near Gorbitsa village as the prospective topotype of this taxon. It was stated that the holotype dimensions in Middendorff’s protologue does not correspond to the proportions of the shell pictured in his book (Pl. 27, Figs. 7–8 12), and that this original holotype picture was “digitally corrected according to the measurements of Middendorff”40. However, this statement is not entirely true, because the shell height vs shell length ratio is 0.47 and 0.46 by the original image and by Middendorff’s measurements12, respectively. This difference is too small and seems to reflect rather slightly inaccurate original measurements than the incorrect holotype picture of Unio mongolicus sensu Middendorff.
To retain the original concept of Unio mongolicus sensu Middendorff as a taxon distinct from Nodularia douglasiae, and to secure the stability of nomenclature, we designate the sequenced specimen RMBH biv229_5 labelled “Russia, Primorye Region, downstream of Gladkaya River (42.7065°N, 130.9084°E), 26.x.2016, Bolotov and Vikhrev leg.” as the neotype of this species (Fig. 5A). The reference sequences accession numbers for the neotype are as follows: MH974549 for COI, MK574414 for 16S rRNA, and MK574555 for 28S rRNA. The shell measurements are as follows: shell length 71.7 mm, height 37.6 mm, width 27.3 mm. The neotype is designated in accordance with the conditions specified in Art. 75 of ICZN, because the name-bearing type specimen was lost, and the authors consider that a name-bearing type is necessary to define the nominal taxon objectively and to avoid further speculations on this issue. We designated a specimen from the Gladkaya River as the neotype, because in this sequenced sample (three genes), we found a specimen that is nearly identical externally to the lost Middendorff’s holotype. The COI sequence of the neotype is very similar to that obtained from a specimen collected from the Shilka River, relatively close to the Middendorff’s type locality (uncorrected p-distance = 0.70%). A sequenced sample from the original type locality is not available.
Genus Nodularia Conrad, 1853
=Amurunio Zatravkin & Bogatov, 1987 (type species: Nodularia lebedevi Zatravkin & Starobogatov, 1984; original designation)20,25.
=Magadaninaia Martynov & Chernyshev, 1992 (type species: Nodularia (Magadaninaia) extremalis Martynov & Chernyshev, 1992; original designation)25,58.
Type species: Unio douglasiae Griffith & Pidgeon, 1833 (monotypy).
Diagnosis. This genus is externally similar to Middendorffinaia, but can be distinguished from it by a narrower, elongated shell, an almost straight hinge plate, comparatively straight or slightly convex dorsal margin, and umbonal sculpture with W-shaped, broad ridges in young shells. However, umbonal sculpture is often weakly developed or absent.
Distribution. Nodularia douglasiae is widespread in the Amur and Razdolnaya basins, in several coastal rivers of the Okhotsk Sea drainage basin up to the Ola River just north of the city of Magadan, and in northwestern Sakhalin (Figs. 5E–H, 6H and Table 1). This species has a plethora of taxonomic names introduced for its conchological varieties from different parts of the Russian Far East (Supplementary Note).
Genus Unio Retzius, 1788
=Tumidiana Servain, 1882 (type species: Unio tumidus Retzius, 1788; subsequent designation by Kantor & Sysoev, 2005)25.
=Crassiana Servain, 1882 (type species: Unio crassus Retzius, 1788; subsequent designation by Graf, 2010)25.
Type species: Mya pictorum Linnaeus, 1758 (subsequent designation by Turton, 183125).
Diagnosis. There are no conchologically similar genera in European Russia and the Urals but introduced populations in the Upper Amur Basin9 can be mistaken with Nodularia and Middendorffinaia. Nodularia has a more elongated, comparatively cylindrical shell. Middendorffinaia differs from Unio by a strongly convex hinge plate and more developed pseudocardinal teeth.
Distribution. Three Unio species were recorded from Russia (Figs. 4A–C, 6E,F, and Table 1). This genus is widely distributed in European Russia and Western Urals, with an isolated native population of Unio tumidus in the Irtysh Basin in Western Siberia59 and Kazakhstan (Table 1). There were a few occasional records of Unio from the Ob’-Irtysh Basin since the middle of the 19th century5,60,61,62. Non-native populations of Unio pictorum and U. tumidus are known to occur in the Upper Amur Basin (Lake Kenon) in Transbaikalia9 (Fig. 6F).
Source: Ecology - nature.com
