Samples were collected from the sub-Antarctic to Antarctic islands (Fig. 1, Table 1). Twelve rafting floating kelps were collected on both sides of the PF during the Antarctic expedition of the RRS James Clark Ross in 2016. Two more kelps were collected South of the PF. M. pyrifera was collected on the beach in DI (South Shetland Islands) during the Distantcom-2 Antarctic cruise in February, 2017. D. antarctica fragments were collected on the beach in front of the Spanish station in Livingston Island in February, 2019 during the Bluebio-2 cruise. Samples were photographed and frozen for further identification of the seaweeds and their epibionts. Samples of rafting kelp ranged from 0.5 to 18.1 kg wet weight. The passenger species traveling upon the kelps reached a total of 7534 specimens (538 ± 637 passengers/kelp, within a range from 0 to 2362 per kelp) and were identified to the lowest possible taxonomic level in the laboratory. The entire rafts were sampled for fauna. Identification of seaweed samples was achieved by studying morphological features, as well as histological examination of the thallus.
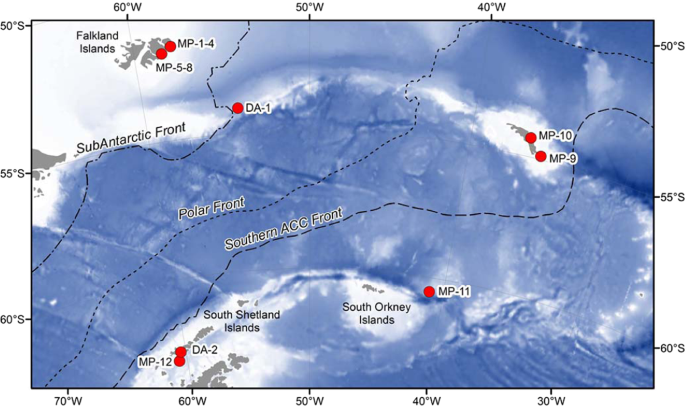
Map of the collecting localities showing the Polar Front (dotted line) and sampling points (in red). DA (Durvillaea antarctica), MP (Macrocystis pyrifera). MP-1–4: Falkland Islands (North), MP-5-8: Mare Harbour (Falkland Islands), DA-1: South of Falkland Islands (Drake passage), MP-9: South Georgia Islands (South), MP-10: South Georgia Islands (North), MP-11: South Sandwich Islands, and MP-12: Deception Island (South Shetland Islands), DA-2: Livingston Island (South Shetland Islands).
Passengers into the cold
Abundance and taxa richness of epibionts found on floating macroalgae in the Southern Ocean vary between the species of kelp (M. pyrifera and D. antarctica) and the individual rafts (Fig. 2). Other rafts, including those formed by D. antarctica, were observed at DI but were not sampled for fauna. Among the four passenger species found alive on M. pyrifera in DI, the most significant in terms of potential ecological impact, other than the non-native kelp itself, is the cheilostome bryozoan Membranipora membranacea. This is a well-known encrusting species with a proven ability to colonise new environments and cause significant damage to ecosystems by limiting the ability of seaweeds to reproduce and grow20. This bryozoan is widely distributed in temperate oceans with distinct populations in the Pacific (North Pacific, Chile, Australia and New Zealand) and Atlantic oceans (North East Atlantic and South Africa) (Fig. 3). M. membranacea has become an established invasive species in the North West Atlantic along the coast of North America and has caused extensive losses of kelp canopy through a process of defoliation21. Although the species is recorded as far north as northern Scandinavia in the Arctic, it has never been previously reported from south of the PF, but it is likely to already be well adapted to cold water conditions, therefore posing more than a hypothetical risk for Antarctic waters.
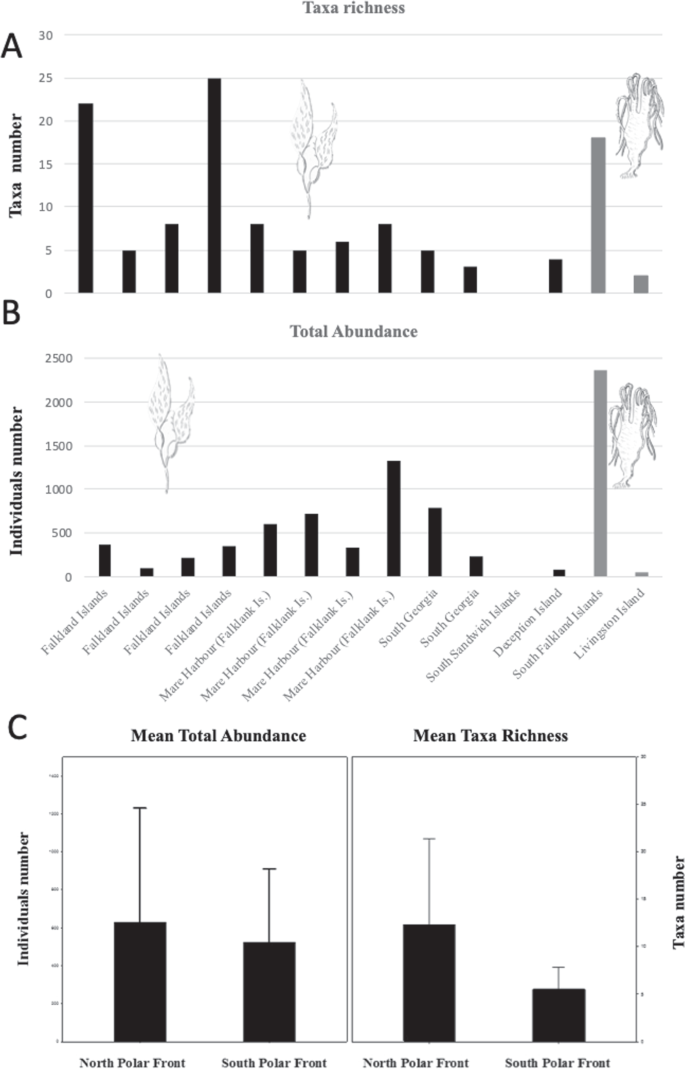
Abundance and taxa richness of epibionts in the rafting algae studied here. Taxa richness (A). Total abundance (B). Black bars: Macrocystis pyrifera, Grey bars: Durvillaea antarctica. Means of total abundance and taxa richness (C) at North and South of the Polar Front (PF) (± s.d.).
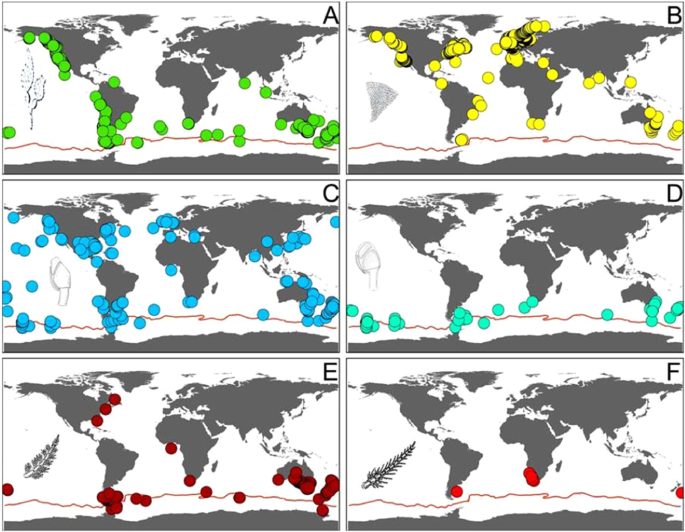
Known distributions of the epibiotic species found associated with Macrocystis pyrifera (A: Distribution of M. pyrifera) on the South Shetland Islands: Membranipora membranacea (B); Lepas anatifera (C); Lepas australis (D); Ballia callitricha (E) and Ballia sertularioides (F). Data from GBIF45.
The combination of having a long-lived planktonic larva (from 2 weeks to 2 months), sexual (hermaphroditic zooids) and asexual reproduction, fast growth rates, effective food acquisition in a wide range of flow rates, ability to form large colonies and to colonize kelps make M. membranacea a successful disperser, colonizer, and invasive species22,23,24. Potentially, these kelp can be transported much farther than bryozoan larvae25,26,27,28,29,30,31. Furthermore, their heavy encrustations may have a negative impact on marine ecosystems by increasing the brittleness of kelp blades, followed by extensive losses of kelp canopy21, and by limiting the ability of the seaweeds to reproduce and grow, specifically interfering with spore release from the kelp blade20. It has also been shown that other species of the same genus may block nutrient uptake and photosynthesis32,33.
The other three species found alive on the kelp in DI have all been previously reported south of the PF. Ballia callitricha and B. sertularioides are shallow water red algae with a general Southern Hemisphere distribution that includes previous records from the Ross Sea, Antarctica (Fig. 3), but not from DI or West Antarctica8. Juveniles and adults of the southern goose barnacle Lepas australis, were also found. This species, commonly found attached to floating substrata such as macroalgae, volcanic pumice, and plastics in the Southern Ocean (Fig. 3), was the only species recently reported on a specimen of giant bull kelp, D. antarctica, found on King George Island, also part to the South Shetland Islands group4.
Using growth rates cited by Fraser et al.4,19 we estimate an age of approximately 30 days for the barnacles, L. australis, found at DI, suggesting that colonization happened in the open sea. Alternative, faster transportation mechanisms may also exist (e.g. shipping vectors and heavy storms). In fact, Lewis et al.34 suggested hull-fouling is likely to be the most important vector for transporting species to Antarctica as ships create novel pathways, moving across currents and often visiting many locations over short periods of time. The increasing ship activity appears to be a very important factor increasing the probability of non–native marine species establishing within the Antarctic region in the coming decades (over 180 ships were active around Antarctica and the sub‐Antarctic islands in 2017–2018, on potentially more than 500 voyages)35. The presence of small–sized specimens of L. anatifera in the kelp found at Livingston Island could also indicate a short-term rafting for this species. Abundant, alive L. anatifera specimens found on D. antarctica fragments in South bay, Livingston Island, represent, in fact, the first Antarctic report for the species, which was described in tropical and subtropical waters of South America36. The potential effects of barnacle colonization in Antarctica are unknown, but in fact, being pelagic rafting species, they seem unlikely to pose any real threat to the shallow water ecosystems, especially as L. australis is already commonly found on rafts and litter in the Southern Ocean. However, their heavy growth could sink the kelp, thus facilitating access to the seafloor for other benthic passengers.
The other floating and beached kelp samples (M. pyrifera and D. antarctica) collected from either side of the PF were found to be carrying organisms within 12 different phyla as passengers (Tables 1 and 2). Each kelp raft examined represented a different, although sometimes overlapping, subset of organisms usually found inhabiting shallow marine habitats. Only one of the floating specimens, MP-11, an example of the non-Antarctic M. pyrifera found near the South Orkney Islands, had no passengers at all. The most commonly found taxa included amphipod crustaceans, polychaete worms, molluscs, and bryozoans (Table 2). The DI floating kelp was the only specimen collected south of the PF carrying M. membranacea, although this bryozoan was frequently found at the Falkland Islands, a potential source of kelp rafts in that region3. Although more studies are needed to know if M. membranacea has become established in the SO, the potential for this species to impact Antarctic ecosystems could be high, not only in DI, as macroalgal substrates are widespread and colder temperatures are not preventing its spread. For example, a recent study based on a baseline data on presence/absence and abundance of this bryozoan near its current northern range limit suggests that the available algal substrate may be more important than temperature in limiting the spread and abundance of M. membranacea37. On the other hand, MP-5, collected from the open ocean north of the PF, was heavily encrusted with thousands of adult and juvenile goose barnacles. This specimen was also host to a rich and varied community of other organisms that are likely to have been associated with the raft before it became dislodged (Table 2).
Other significant findings included the brachyuran crabs Halicarcinus planatus and Peltariom spinulosum in the M. pyrifera fragments washed up on the shore at the Falkland Islands. H. planatus was first recorded in Antarctica at the shores of the South Orkney Islands in 190338. It was reported again by Aronson et al.39 at the external side of the caldera of DI, supporting the hypothesis that DI could be the entrance gate for non-native species. H. planatus is a widely-distributed species in temperate waters, found from New Zealand to the Falklands and southern South America, as far north as Peru and Argentina40,41. H. planatus has also been found alive on floating kelp42. Although H. planatus was not found in our previous studies at Deception and Livingston Islands8, we did find it on M. pyrifera washed up on the shore of the Falkland Islands (pers data 2016, SO-AntEco expedition), which could easily be re-floated by high tides or rough weather. The impact of these crabs on local species is not known but could potentially be devastating due to the absence of durophagous fauna in Antarctic shallow benthic ecosystems43,44.
Rafting to the south
The transport of organisms on ships’ hulls or in ballast water can take less than 4% of the time it would take to reach the same destination by rafting11. Although this significant reduction in time taken to reach Antarctica might allow a wider range of species to reach the continent alive, they would still need to be capable of surviving the conditions at their destination in order to become established. As such, our observation of a species with a documented track record of invasive and negative ecological impacts, such as Membranipora membranacea, in an active volcano (DI), with warmer, more favourable conditions, is very significant. The species reported here are common and well-distributed organisms and thus have the potential to persist or even thrive in the milder conditions of the caldera of DI (Fig. 4). It could only be a matter of time before some of these species acclimatize to the Antarctic environment and spread. These findings are even more relevant in the current context of global change, which could facilitate the survival of these species in other Antarctic environments once settled in favourable areas, such as DI, further reaching other places around the Antarctic peninsula. Therefore, these species may be useful indicators of climate change in Antarctic habitats and should be carefully monitored during the next years.

Rafting kelp and passengers. Macrocystis pyrifera (A) with passengers found at Deception Island, and Durvillaea antarctica (B) with cirripeda from Livingston Island, South Shetland Islands, Antarctica. (C–F) Passengers found on M. pyrifera at DI: the red alga Ballia callitricha (C); the bryozoan Membranipora membranacea (D); the cirripeda Lepas australis (E); and the cirripeda L. anatifera (F) on D. antarctica from Livingston Island.
Source: Ecology - nature.com
