Seafloor mapping
The Neopycnodonte bioconstructions were studied in two different areas of the southern Apulian coast, Otranto (OT) and Santa Maria di Leuca (SML) (Fig. 1). In the OT area, Neopycnodonte bioconstructions were discontinuously detected along 600 m of the coastline within a bathymetric range of 45–64 m, reaching a total length of 200 m (Fig. 2). In the SML area, a carbonate formation built by the mollusc bivalve almost uniformly covered the northern and eastern sides of the cliff for a total length of approximately 450 m in the bathymetric range of 45–70 m (Fig. 3).
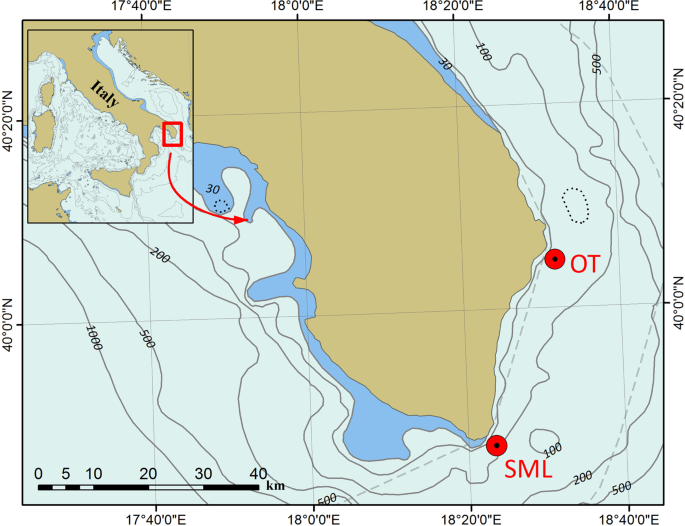
Locations of the two study areas: Otranto (OT) and Santa Maria di Leuca (SML). They occur off the southern Italian coast and are identified by red circles. Map has been created with ESRI ARCMAP 10.2, available at https://support.esri.com/en/products/desktop/arcgis-desktop/arcmap/10–2–2.
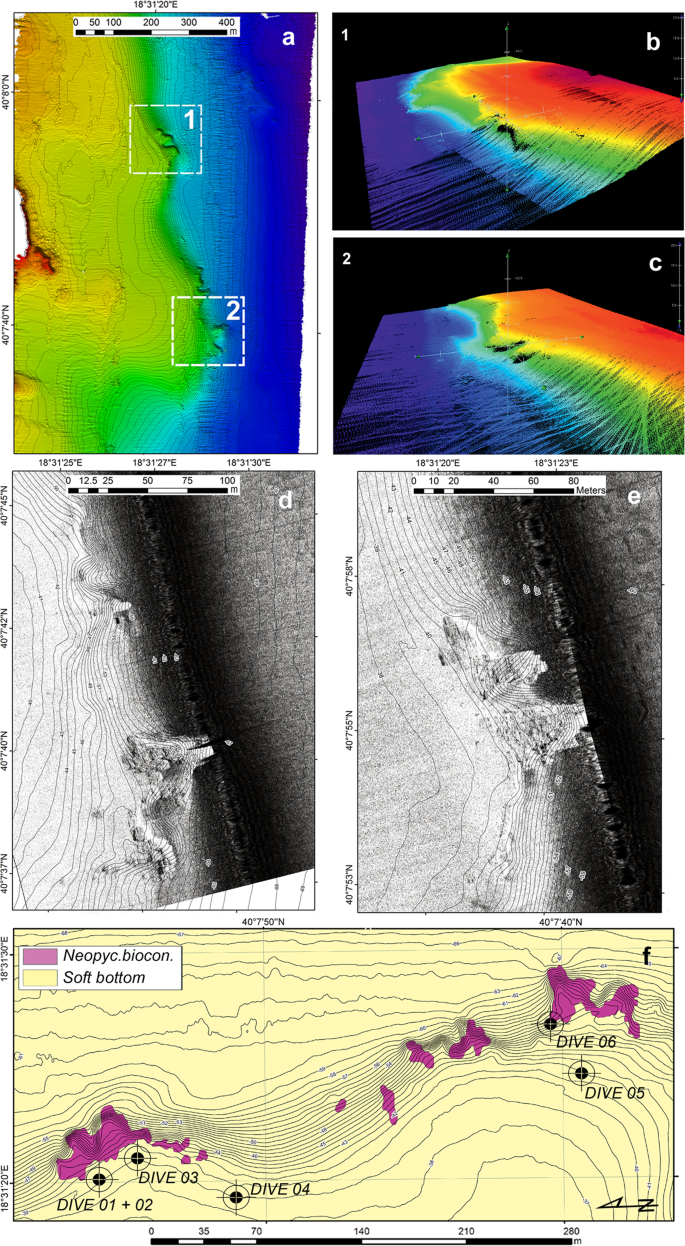
Geophysical survey of the Otranto area and localization of dive points. (a) Digital elevation model of the seafloor derived by the multibeam survey. (b,c) Areas 1 and 2 are characterized by the largest slope gradient. (d,e) Details of the raw data from the Side-Scan Sonar survey in areas 1 and 2. Note the presence of hard substrate along the slope. (f) Classification of the seafloor and localization of dive points (black circles). Maps have been created with: (a) ESRI ARCMAP 10.2 + DTM and image produced by CARIS HIPS 9; (b,c) CARIS HIPS 9 (Subset editor); (d,e) ESRI ARCMAP 10.2 + SSS mosaics produced by CARIS SIPS 9; (f) ESRI ARCMAP 10.2, all available at https://support.esri.com/en/products/desktop/arcgis-desktop/arcmap/10–2–2 and https://www.teledynecaris.com/en/products/hips-and-sips/.

Geophysical survey of the Santa Maria di Leuca area and localization of dive points. (a) Digital elevation model of the seafloor derived by the multibeam survey. (b) Details of the ESE-WNW-oriented slope. Note that the slope seems to be very irregular along the portion with a higher gradient. (c) Raw data from the Side-Scan Sonar survey. (d) Classification of the seafloor and localization of dive points (white circles); the bioconstructions can be easily mapped, while shallower and deeper sectors seem to have a similar, even though unclear, acoustic signal. Maps have been created with: (a) ESRI ARCMAP 10.2 + DTM and image produced by CARIS HIPS 9; (b) CARIS HIPS 9 (Subset editor); (c) ESRI ARCMAP 10.2 + SSS mosaics produced by CARIS SIPS 9; (d) ESRI ARCMAP 10.2, all available at https://support.esri.com/en/products/desktop/arcgis-desktop/arcmap/10–2–2 and https://www.teledynecaris.com/en/products/hips-and-sips/.
The two study areas, OT and SML, showed similar geomorphological features (Figs. 2 and 3). The seafloor geometry was dominated by the presence of a slope that connected a large coastal flat area to deeper sectors (depth range: 39–64 m in OT and 27–70 m in SML). This slope ran parallel to the coastline in the OT area (NNW-SSE, Fig. 2a), while in the SML area, it ran along an ESE-WNW direction, transversally to the coastline (Fig. 3a). In both cases, the slope was locally steep and showed irregular morphology (Figs. 2b,c and 3b).
Data from the Side-Scan Sonar (SSS) survey allowed a more detailed assessment of the nature of the seafloor. In the OT area, the acoustic signal was monotonous in the shallower (39 m) and deeper sectors (64 m), likely in relation to the presence of soft bottom (Fig. 2d,e). Rocky substrate randomly occurred in only the slope sectors, forming localized submerged “headlands” with an irregular slope and a nearly flat top.
In the SML area, the flat and shallow sectors (25–27 m) had a rocky substrate (Fig. 3c). The mosaicked acoustic signal suggested the presence of coralligenous bioconstructions or concretions, while close to the top of the slope (30 m), the substrate showed acoustic features that can be interpreted as small bioconstructions with irregular morphology (Fig. 3c,d). The slope was stiff and continuous, showing a complex acoustic signal likely related to the presence of large-scale bioconstructions (comparable with the irregular slope of the multibeam dataset (Fig. 3b). Deeper areas (70 m) seemed to have a fine-grained sediment substrate.
Structure of the bioconstructions: building and structuring taxa
In both study areas, the bioconstructions developed in thick pinnacles or globose formations, protruding perpendicularly with respect to the cliff for approximately 50 cm at OT and for more than 1.5 m at SML and often interconnected with one another to form a framework of high structural complexity (Fig. 4a,b). The pinnacles were organized in successive terraces proceeding from the top to the bottom of the bioconstruction (see Supplementary Videos 1 and 2). According to the analysis of the video images, sampled material and resin slices, the pinnacles and their basal layer were mainly formed by the massive, multilayered aggregation of shells of Neopycnodonte cochlear (Poli, 1795) (Figs. 4c and 5). In both study areas, N. cochlear occurred in 100% of the analysed images, with average covering values of 84 and 82% in OT and SML, respectively (Fig. 6). At both sites, most of the bioconstructions were composed of dead specimens of N. cochlear. Live specimens were present in scattered clusters of a few individuals (6–20) grafted onto the superficial layer of the bioconstruction. Specifically, the large-scale analysis of the resin slices showed how the general framework of the bioconstruction was always shell supported and derived from the complex superposition of new valves on the preceding ones (Fig. 7). In general, N. cochlear specimens seemed to be irregularly arranged with respect to each other (Fig. 5), with individuals settling on the surface of older shells and sharing few points of contact with adjacent valves (Fig. 7a). Locally, some shells were arranged parallel to each other, thus increasing the contact surface between adjacent valves (Fig. 7b,c). The random orientation of the shells and the presence of point-like contacts determined the formation of an overall porous structure. At the same time, the contacts between larger surfaces favored the stability of the bioconstruction.

The Neopycnodonte bioconstructions. Underwater images of the bioconstructions: (a) globose formations at Otranto and (b) thick pinnacles at Santa Maria di Leuca protruding perpendicularly with respect to the cliff and interconnected with one another to form a framework of high structural complexity. (c) Detail of the Neopycnodonte bioconstruction formed by the massive, multilayered aggregation of shells. (d, e) Scleractinian facies. (e) Detail showing the main structuring taxa: Cladopsammia rolandi/Leptopsammia pruvoti complex (yellow corallites) and Hoplangia durotrix (light brown corallites). (f) Corallium rubrum facies. (g) Large portion of the substrate covered by the encrusting sponge Dendroxea lenis (grey). (h) Paramuricea clavata facies characterized by large colonies. (i, j) Detail of Neopycnodonte bioconstruction heavily infested by the boring sponge Siphonodictyon infestum (arrows) in a fresh sample (i) and in a sample embedded in resin (j).

Large sample of the bioconstruction after collection. Neopycnodonte shells mainly appear irregularly arranged, forming a framework of high structural complexity.
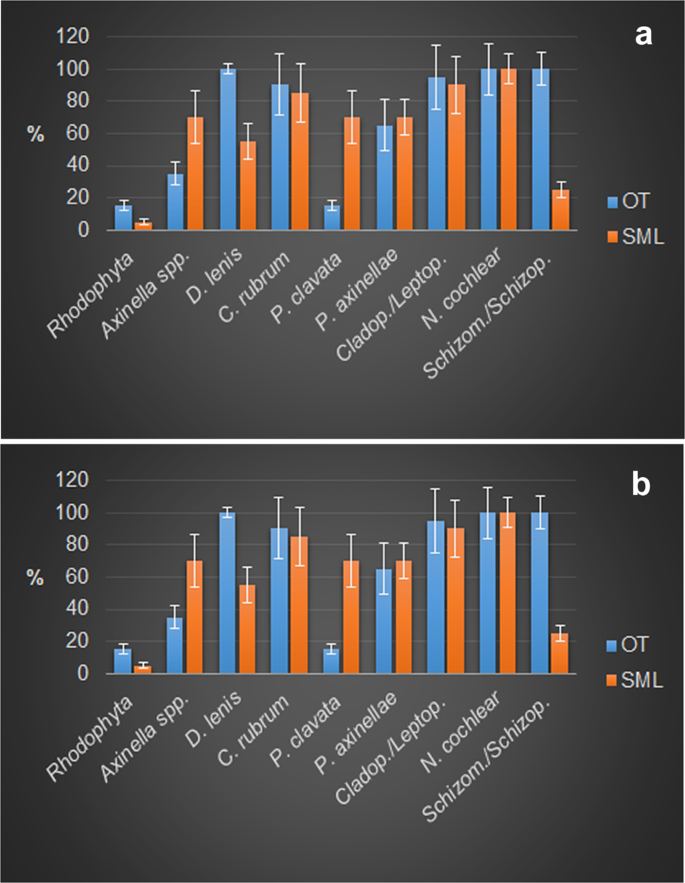
Contribution of the different structuring taxa to the bioconstruction at Otranto (OT) and Santa Maria di Leuca (SML). (a) Frequency and (b) covering values of the main structuring taxa (mean % ± s.e.). (D. = Dendroxea; C. = Corallium; P. = Paramuricea; P. = Parazoanthus; Cladop./Leptop. = Cladopsammia/Leptopsammia; N. = Neopycnodonte; Schizom./Schizop. = Schizomavella/Schizoporella).
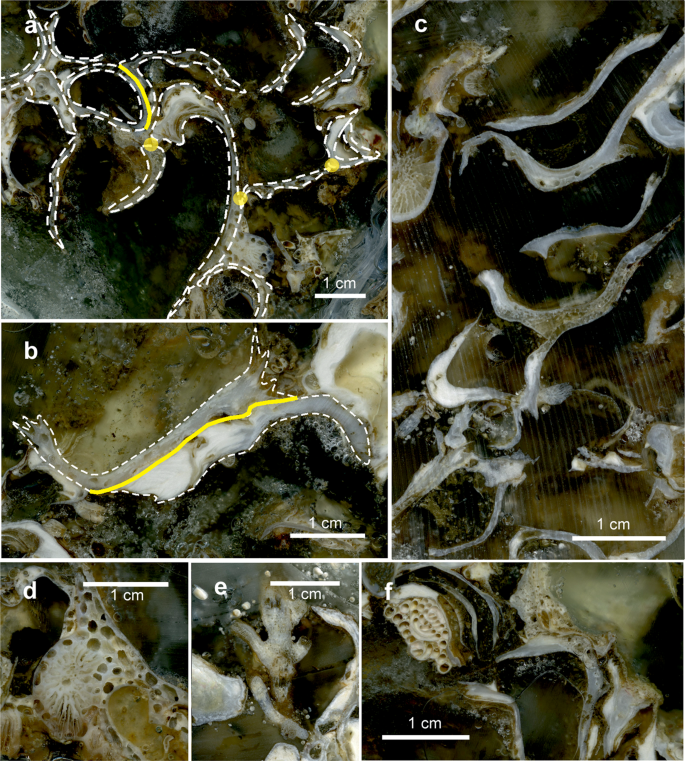
Details of the large-scale slices of the bioconstruction impregnated with epoxy resin. (a) Shells sharing single points (yellow circles) or large surfaces (yellow line). (b) Parallel Neopycnodonte shells that grow together, sharing large surfaces (yellow line). (c) Locally, the bioconstruction shows a remarkable porosity. (d) Traces of sponge perforations. (e) Scleractinian corallites on a mollusc shell. (f) Serpulids and bryozoans encrusting the surface of Neopycnodonte shells.
Among the other taxa structuring the bioconstructions, there were cnidarians, serpulids and bryozoans, all contributing to the deposition of calcium carbonate, and poriferans, helping to bind shells together or eroding carbonate by boring species. Indeed, boring sponges were often recorded living into the carbonate structures (Figs. 4i,j and 7d), and colonial scleractinians and serpulids were found within the concretion (Fig. 7e,f). In particular, among the secondary structuring taxa, the scleractinians Cladopsammia rolandi Lacaze-Duthiers, 1897, Leptopsammia pruvoti Lacaze-Duthiers, 1897, and, to a lesser extent, Caryophyllia (Caryophyllia) inornata (Duncan, 1878) and Hoplangia durotrix Gosse, 1860, were the most frequent structuring species (Fig. 4d,e and 6), having a constant presence across the bioconstructions. The alcyonacean Corallium rubrum (Linnaeus, 1758) also strongly contributed to the bioconstructions (Fig. 6). It showed a patchy distribution, with aggregates of several specimens concentrated below the pinnacles of the structure (Fig. 4f).
The outer portions of the bioconstructions as well as the reef interstices were extensively encrusted by serpulid tubes. In particular, most spirorbid polychaetes, especially Protolaeospira (Protolaeospira) striata (Quiévreux, 1963), Pileolaria militaris Claparède, 1870 and Vinearia koehleri (Caullery & Mesnil, 1897), colonized the bare parts of the substrate, such as the external edges of the shell of living Neopycnodonte specimens and their smooth inner parts, corresponding to the pioneering role that these organisms played in the community colonization pattern. Other spirorbid species exhibited their particular adaptation to the cryptic and dark crevices of the bioconstruction according to their small dimensions and often-wrapped tubes.
The bryozoans Schizomavella spp. and Schizoporella spp., particularly well represented in the OT study area (Fig. 6), formed thin crusts on the reef surface that contributed to the compactness of the structure. Sponges were mainly represented by encrusting species covering large portions of substrate. Among them, Dendroxea lenis (Topsent, 1892) occurred frequently in both study areas, with covering values that reached 42% of the substrate of the bioconstruction (Figs. 4g and 6). In contrast, massive and erect forms were less represented in both study areas, with the exception of small specimens of Axinella spp. (Fig. 6), which were mainly concentrated in the sub-horizontal portions of the substrate. Among the boring sponges, Siphonodictyon infestum (Johnson, 1889) played an important role as a bioeroder of the bioconstruction (Fig. 4i,j).
In addition, the reef’s crevices were also inhabited by Hiatella spp. molluscs living as nestlers or as borers, enlarging the reef’s holes. Additionally, the soft-bottom bivalve Kellia suborbicularis (Montagu, 1803) preferred the sediment trapped in the crevices and the spaces among the Neopycnodonte shells. Finally, red coralline algae were only sporadically detectable during the analysis of both images and biological samples (Fig. 6).
Regarding large epibenthic taxa, dense populations of the gorgonians Paramuricea clavata (Risso, 1826) and Eunicella cavolini (Koch, 1887) characterized the seascapes of both the habitats at both sites, representing the main 3D habitat makers, although at OT gorgonians were limited to a few areas of the bioconstruction (Fig. 4h and 6).
The mapping of the area occupied by different taxa in the large-scale slices of the bioconstruction (Fig. 8) showed the following average percentage values: Neopycnodonte shells: 73.8 ± 7.7%, scleractinians: 13.6 ± 10.2%, serpulids: 8.8 ± 7.6%, bryozoans: 2.3 ± 1.2%, encrusting algae: 1.3% ± 1.2% (Fig. 8c). The analysis of images showed that the bioconstructions were characterized by marked porosity (73.2 ± 3.3%) that was due to spaces within and between individuals and small-scale porosity related to bioerosion (Figs. 8 and 9). The boring sponges occurred in all sampled material at both sites, where they heavily bioeroded carbonate structures, showing a clear decreasing gradient of perforation from the oldest to the youngest parts of the bioconstruction (Fig. 9).
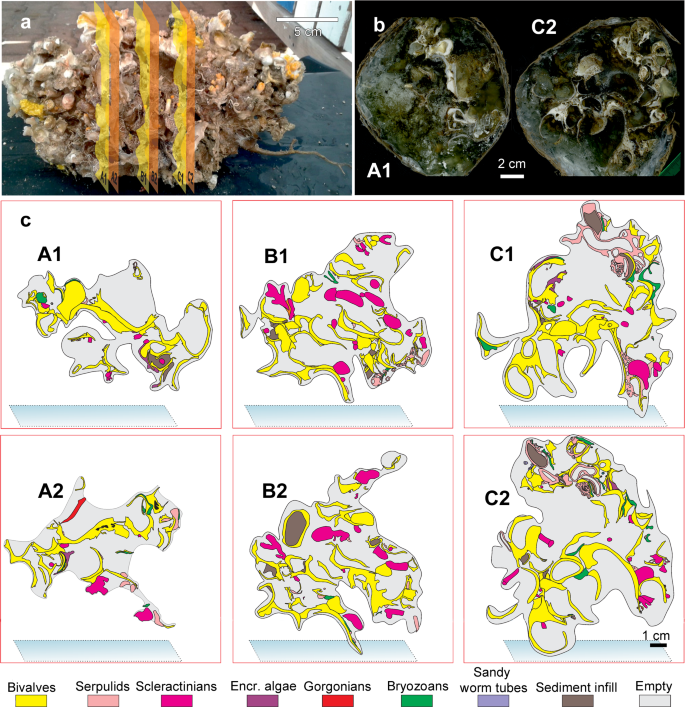
Bioconstruction block with details of the main structuring taxa identified. (a) Sampled block with indications of cut planes (yellow and orange polygons). (b) Examples of high-resolution images of the large-scale slices of the bioconstruction. (c) Compositional map of the different taxa.
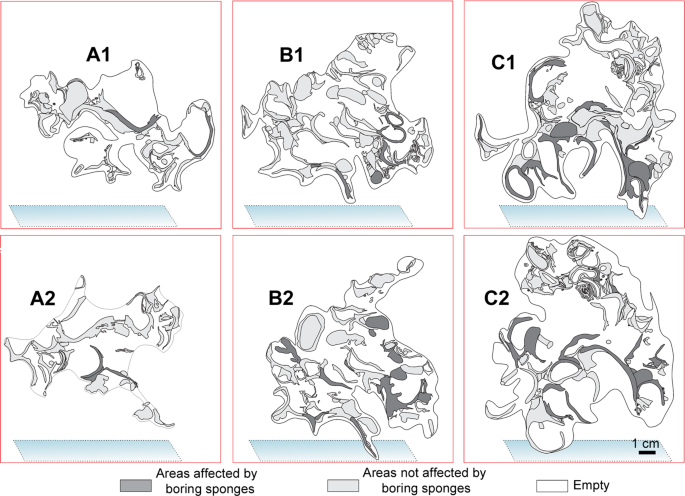
Distribution of the perforations produced by boring sponges in the same slices of Fig. 8.
Taxonomic accounts and autecological features
Analysis of the biological samples revealed the presence of a total of 165 structuring taxa (153 identified to the species level), 110 of which were detected at OT and 136 at SML (Supplementary Table S1), and 81 (49%) were shared by both sites. Overall, the phylum Porifera had the greatest species richness (65 taxa), followed by Annelida (38 taxa) and Bryozoa (34 taxa), while algae were present to a lesser extent (5 taxa). The patterns of species of the different taxa showed total overlap between the two study sites for algae, cnidarians and bivalves, with SML hosting all the taxa recorded at OT plus some exclusive ones. In contrast, sponges, annelids and bryozoans diverged in terms of species composition (Fig. 10).
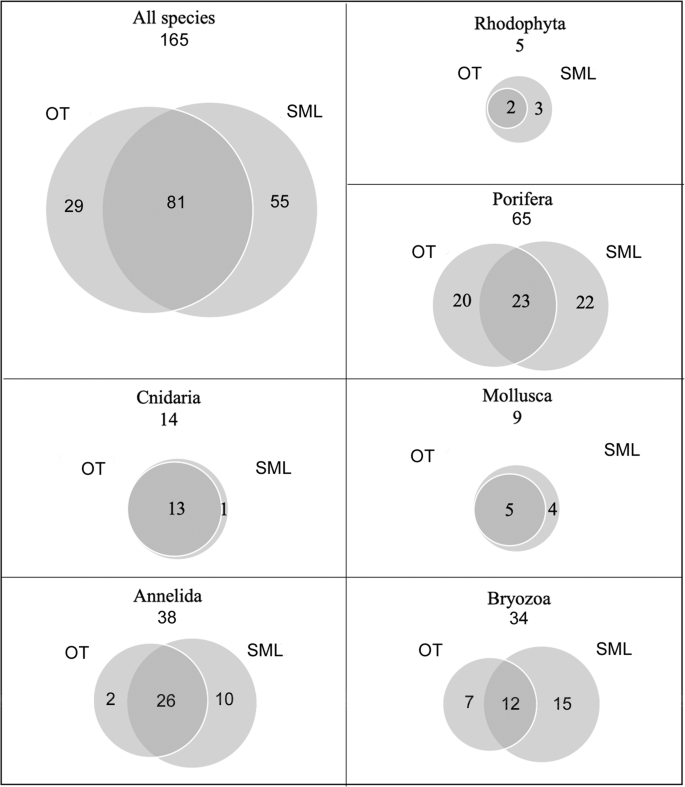
Venn diagrams showing relationships among the sampling areas at Otranto (OT) and Santa Maria di Leuca (SML) in terms of total species richness and main structuring taxa. The numbers in the overlapping areas indicate the shared taxa, those in the external areas indicate the exclusive taxa.
Algae
A total of 5 species of the class Florideophyceae were identified (Supplementary Table S1). Three of them are non-geniculated encrusting forms: Titanoderma pustulatum (J.V. Lamouroux) Nägeli, 1858 and Lithophyllum stictiforme (J.E. Areschoug) Hauck, 1877 belonging to the order Corallinales, and Mesophyllum alternans (Foslie) Cabioch & M.L. Mendoza, 1998 belonging to the order Hapalidiales. Jania sp. and Amphiroa sp. are geniculated species belonging to the order Corallinales. T. pustulatum and M. alternans are the only species that were detected in both study areas. Coralline species showed a patchy pattern in both study areas, where they were represented by small thalli, with a maximum surface covering of a few square centimetres. Encrusting species were attached to tiny rocks, scleractinians and mollusc shells. Geniculated species were attached to encrusting coralline species or other Rhodophyta species.
Porifera
Porifera encompassed 65 taxa, with 61 identified to the species level, as follows: 4 species of Homoscleromorpha (1 order, 2 families, 4 genera) and 61 of Demospongiae (3 subclasses, 13 orders, 30 families, 41 genera) (Supplementary Table S1). The order Dictyoceratida was most represented, with 4 families, 6 genera and 11 species. Poecilosclerida was represented by 4 families, 5 genera and 8 species. Finally, Axinellida, Haplosclerida, and Tetractinellida were other representative orders. Most of the sponge taxa recorded show an Atlanto-Mediterranean distribution, and 12 taxa are currently reported only in the Mediterranean Sea28.
The number of sponge taxa found at the study sites was 43 at OT and 45 at SML, 23 of which were shared, with a high number of exclusive species (20 and 22, respectively) (Fig. 10).
In general, encrusting forms prevailed at both sites, and the few massive and erect species, when present, were of small size. Among the encrusting forms, Dendroxea lenis (Topsent, 1892) showed the highest frequency (Fig. 6a), Axinella verrucosa (Esper, 1794) was the most frequent sponge among erect species (Fig. 6), and massive species were only sporadically detected. Seven species were boring sponges, but only one of them (Siphonodictyon infestum (Johnson, 1889)) was widely represented in samples from both sites; this species is able to produce large boring chambers in the shell walls of N. cochlear (Fig. 4i,j).
Cnidaria
Fourteen anthozoan taxa belonging to the orders Alcyonacea (3 families and 4 genera), Scleractinia (4 families, 9 genera) and Zoantharia (1 family, 1 genus) were detected in both study areas (Supplementary Table S1). Most of the species recorded have an Atlanto-Mediterranean distribution, with the exception of Eunicella cavolini (von Koch, 1887) and Parazoanthus axinellae (Schmidt, 1862), whose current known distribution is limited to the Mediterranean29.
The two study areas showed high similarity in terms of composition of the Anthozoa. Indeed, with the exception of Alcyonium coralloides (Pallas, 1766), detected at SML only, all the species were found at both sites (Fig. 10).
The order Scleractinia was the most represented in terms of species richness and frequency of occurrence. Scleractinians colonized the shells of N. cochlear or settled inside and between the valves of the empty shells throughout the bioconstruction. Hoplangia durotrix Gosse, 1860 and Caryophyllia (Caryophyllia) inornata (Duncan, 1878) were particularly abundant, scattered throughout the framework. Cladopsammia rolandi Lacaze-Duthiers, 1897 and Leptopsammia pruvoti Lacaze-Duthiers, 1897 formed large facies mainly in shaded regions of the bioconstruction (Fig. 4d,e). Finally, C. (C.) smithii Stokes & Broderip, 1828, Monomyces pygmaea (Risso, 1826) and Stenocyathus vermiformis (Pourtalès, 1868) were more rarely recorded.
Among the alcyonaceans, Paramuricea clavata (Risso, 1826) showed the highest density, with large colonies reaching 50 cm in fan diameter (Fig. 4h).
Mollusca
The living mollusc fauna sampled at OT and SML was represented by 9 species belonging to the classes Gastropoda (5) and Bivalvia (4) (Supplementary Table S1). Among the Gastropoda, 4 species belonged to the order Littorinimorpha and 1 to the order Lepetellida. With regard to bivalves, N. cochlear belonged to the order Ostreida and to the family Gryphaeidae, and the other 3 species belonged to the orders Galeommatida (1 species) and Adapedonta (2 species). All the mollusc species have an Atlanto-Mediterranean distribution, with the exception of the gastropods Alvania carinata (da Costa, 1778) and Sandalia triticea (Lamarck, 1810), which have a distribution limited to the eastern Mediterranean basin (Horton et al., 2019). The two study areas shared 55% (2 Gastropoda and 3 Bivalvia) of the mollusc fauna, while the remaining 45% (4 species) was exclusively found at SML (Fig. 10). In general, the mollusc fauna was characterized by low abundance values, with the exception of N. cochlear, the main builder of the bioconstructions. Living specimens of N. cochlear formed clusters of a few individuals scattered on the upper part of the calcareous framework, while most of the bioconstruction was composed of dead specimens. The abundance of living specimens of N. cochlear calculated for 300 ml of the bioconstruction varied from 1.7 ± 0.7 to 9.1 ± 2.4 at OT and SML, respectively. In particular, the living specimens detected in the OT samples were mainly represented by juveniles (1.3 ± 0.2%) with the length of the main valve measuring approximately 1 cm.
Regarding the other mollusc species, most were recorded inside and between the valves of dead specimens of N. cochlear, although Vermetus triquetrus Bivona-Bernardi, 1832 colonized the exposed surface of the bioconstruction, covering some of the Neopycnodonte valves. The gastropod Pseudosimnia carnea (Poiret, 1789) is locally abundant and was mainly found to be associated with red coral colonies, on which it is parasitic.
Annelida
Thirty-eight species of Serpulidae, with 29 Serpulinae and 9 Spirorbinae, were recorded (Supplementary Table S1). Almost all species (36) were collected at SML, and 28 species were found at OT, with 26 shared between the two sites. Most species were found with few individuals, while Vermiliopsis infundibulum (Philippi, 1844), Semivermilia crenata (O.G. Costa, 1861), Filogranula annulata (O.G. Costa, 1861) and Semivermilia pomatostegoides (Zibrowius, 1969), together with some spirorbids, such as Protolaeospira (Protolaeospira) striata (Quiévreux, 1963) and Pileolaria militaris Claparède, 1870, were particularly abundant. Such polychaetes have a Mediterranean and north-eastern Atlantic distribution, and some of them are cosmopolitan, e.g., V. infundibulum, S. vermicularis, Josephella marenzelleri Caullery & Mesnil, 1896, P. militaris, Neodexiospira pseudocorrugata (Bush, 1905), and Janua heterostropha (Montagu, 1803); 6 species are endemic to the Mediterranean, i.e., Placostegus crystallinus (non Scacchi, 1836) sensu Zibrowius, 1968, S. pomatostegoides, V. monodiscus Zibrowius, 1968, and Spirobranchus lima (Grube, 1862), and 3 species, i.e., Serpula cavernicola Fassari & Mollica, 1991, S. annularis Dillwyn, 1817 and Nidificaria clavus (Harris, 1968), only occur in the Mediterranean Sea, Gibraltar area and Canary Islands. The polychaete species exhibit a high level of adaptive radiation and can be ascribed to different ecological groups30,31,32: meso-infralittoral shelf species, e.g., Spirobranchus polytrema (Philippi, 1844), S. triqueter (Linnaeus, 1758) and Janua heterostropha (Montagu, 1803); characteristic coralligenous species, e.g., S. crenata and Vinearia koehleri (Caullery & Mesnil, 1897); detrital continental shelf species, e.g., S. cribrata (O.G. Costa, 1861) and Spirorbis (Spirorbis) cuneatus Gee, 1964; deep-water and bathyal species, e.g., V. monodiscus, Serpula israelitica Amoureux, 1977, and Filogranula gracilis Langerhans, 1884; and cave species, e.g., S. cavernicola and F. annulata.
Bryozoa
Thirty-four taxa of bryozoans were identified: 30 belonging to the class Gymnolaemata, order Ctenostomatida, and 4 belonging to the class Stenolaemata, order Cyclostomatida (the latter were not identified to the species level) (Supplementary Table S1). The bioconstructions at SML showed a higher species richness (27 taxa) with respect to that at OT (19 taxa). Twelve taxa were shared between the two sites, while the exclusive species accounted for 21% at OT and 44% at SML (Fig. 10). Most of the bryozoan species are distributed in the north-eastern Atlantic Ocean and largely in the Mediterranean Sea, but some of them, e.g., Schizoporella mutabilis Calvet, 1927, Schizoretepora serratimargo (Hincks, 1886), Rhynchozoon sp., and Pentapora fascialis (Pallas, 1766), are endemic to the Mediterranean.
Many taxa were found living close to one another. Most of them exhibited both thick (11 taxa) and thin (7 taxa) encrusting habitus, others were present in petraliform and celleporiform colonies (3 species), and 7 taxa were found to form erect colonies. The encrusting species of the genus Puellina were the major occupiers of the substrate, together with Schizomavella and Schizoporella spp., which developed sheets that extensively covered the surface of the bioconstruction. Moreover, encrusting bryozoans grew epibiotically on serpulid tubes and on other bryozoan colonies. Bryozoans of the genera Crassimarginatella and Beania, with petraliform colonies, and those of the genera Rhynchozoon and Turbicellepora, with celleporiform colonies, populated interstices, cavities and crevices of the bioconstruction. The few erect bryozoans mainly colonized the outer edges of Neopycnodonte shells with both rigid (Myriapora truncata (Pallas, 1766) and Crisia sp.) and flexible (Bugula gautieri Ryland, 1962) colonies.
Source: Ecology - nature.com
