Application of three previously reported, popular CHD1-targeting primer sets for molecular sexing of the Japanese murrelet, and preparation of a new Japanese murrelet species-specific primer set
The three frequently used CHD1-targeting primer sets were applied for molecular sexing in the Japanese murrelet. Two of these three primer sets, 2250 F/2718 R and CDH1F/CDH1R, produced electrophoretic patterns similar to those reported previously in many species of birds. Specifically, males yielded a single band, while females yielded two bands, one of which matched the size of the male-specific band. The shared band presumably corresponded to a CHD1 homolog on chromosome Z, while the second band appeared to correspond to the CHD1gene on chromosome W. In contrast, the P2/P8 primer set yielded a single band from all animals; presumably this band corresponded to a mix of CHD1Z and CHD1W amplicons running at the same size or this primer set did not permit amplification from chromosome W. However, comparison of the electrophoretic patterns indicated there were noteworthy differences in the sizes of the products. When molecular sexing is carried out using non-invasive samples from wild birds, smaller-sized fragments seem to be amplified more efficiently than are larger-sized fragments. Therefore, we designed a new Japanese murrelet-specific primer pair predicted to yield a smaller PCR product from chromosomes Z and W than were obtained from the previously described primer sets. Indeed, the fragments amplified with the newly designed primer set were smaller in size and more efficiently amplified than those obtained with the previously reported, popular primer sets 2250F/2718R and CDH1F/CDH1R.
Estimation of the utility of the species-specific primer set for Japanese murrelet
Because of many restrictions on the capture of specific birds and on the collection of invasive samples, it was not easy to obtain enough samples as controls. Furthermore, the Japanese murrelet is a monomorphic species, so it is difficult to distinguish female from male based solely on external morphological survey; this point means that it is hard to rapidly and accurately determine animal sex in the field. Therefore, the species-specific primer set CHD1F1/CHD1R1, which was generated as part of the present study, was evaluated for its utility in accurate molecular determination of sex in the Japanese murrelet based on the following reasoning. First, comparative tests were performed using three of the primer sets described above (CHD1F/CHD1R, 2250F/2718R and CHD1F1/CHD1R1) along with the DNA samples extracted from blood clots of Japanese murrelet selected at random, without prior information about animal sex. The results obtained with each primer set were found to be in complete agreement with each other. Second, the newly designed primers for the Japanese murrelet corresponded to DNA sequences homologous to CHD coding regions highly conserved among many avian species. The highly homologous regions in the primers were expected to make it possible to use this primer set for molecular sexing in a variety of bird species. As a practical example, the primer set was applied to another bird species, Japanese quail, which presents sexual dimorphism as adults and in which sex is readily identified based on external morphological features. Within the Japanese quail, there was no discrepancy between molecular sexing with the species-specific primer set and sexing by traditional methods based on morphology (Supplementary Fig. 2). Third, one Japanese murrelet was found dead in Biroudo island, permitting direct determination of sex by inspection of reproductive organs at necropsy. In parallel, a DNA sample was extracted from the feathers of this bird, permitting correlation of the sex as determined by molecular sexing with the CHD1F1/CHD1R1 primer set, and sex as determined by inspection at necropsy. Although there were not additional Japanese murrelet samples for which sex was known both by molecular sexing and by anatomical structures, the existing samples worked well as a positive control, permitting a distinction between results from males and females. Therefore, we conclude that the new primer set provides accurate results when used for sexing of the Japanese murrelet.
A variable number of tandem repeats serve as a DNA marker on chromosome W of Japanese murrelet
When CHD-based molecular sexing was carried out using the species-specific primer set, CHD1F1/CHD1R1, and the products were separated by agarose gel electrophoresis, two different patterns were observed in female samples (Fig. 1, Supplementary Figs. 1 and 2), in which the patterns were clearly differentiated by the difference of space in width between the common (Z-derived) band shared between males and females and two patterns obtained from female Japanese murrelets reflect a polymorphism within this species. As noted above, agarose gel electrophoresis of the products from males and females yielded distinct patterns: two bands were observed in females compared to one in males. The upper fragment was shared in common by both male and female samples and therefore was presumed to be derived from the CHD-Z allele. Additionally, a female-specific (presumably W-derived) band was detected. However, female Japanese murrelets yielded either of two different sizes of W-derived amplicons, defined as long-type or short-type, on agarose gel electrophoresis (Fig. 1, F1 and F2; Supplementary Fig. 1, ZWL and ZWS; Supplementary Fig. 2, Lanes 5 and 6). PCR products from female short-type (WS) and long-type (WL) of chromosome W were cloned into T-vector and analyzed by polyacrylamide gel electrophoresis (Supplementary Fig. 1). The resulting fragments were subjected to sequencing (Fig. 2); an alignment between the sequences is shown schematically in Fig. 3 (MN519214 from CHD1Z gene and MN519215 from CHD1W gene). Alignment of the two different DNA sequences indicated that the polymorphism observed in chromosome W is derived from variation in the number of 32-bp-repeat units. Specifically, WL has two repeat units. The DNA sequence alignments also revealed that each of the three PCR products obtained from the Japanese murrelet using the CHD1-F1/R1 primer set consist of a CHD intron flanked by pieces of the two adjacent exons. The intron starts with a GT and finishes with an AG, which appears to be in accordance with the rule for exon-intron junctions. The lengths of the DNA fragments of the intronic regions for the Z, WL, and WS-derived fragments are 312 bp, 203 bp and 171 bp, respectively.
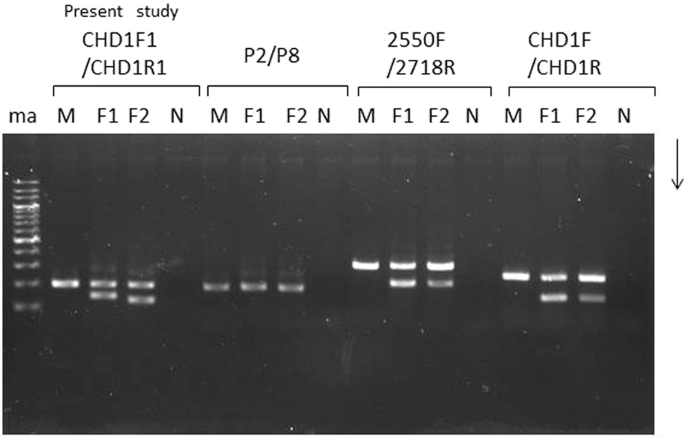
Comparison of agarose gel electrophoretic patterns of the PCR products amplified by each of the four primer set tested for sexing of Japanese murrelets. CHD1F1/CHD1R1 was designed in the present study; P2/P8, 2550F/2718R, CHD1F/CHD1R were described previously. For each primer set, three samples (consisting of one male (M) and two female(F1 and F2) Japanese murrelets were analyzed. N indicates the negative controls (reactions performed without sample DNA). With the exception of P2/P8, all primer sets were able to distinguish the chromosome Z-derived fragments from the smaller chromosome W-derived fragments. The ma line indicates the 200-bp DNA ladeder for a marker DNA.

Alignment of three DNA sequences of Z chromosome (Z/Z male), W chromosome L-type (clone WL) and W chromosome S-type(clone WS), which are GHD1-related PCR products including the region of the parts of two exons and the intron between the exons. Dash(–) indicates the delete of nucleotide. The intron starts at GT and finished at AG, this sequence agrees with GT-AG rule. The boxed DNA sequences indicate the primer sequences. Comparison of DNA sequences among the PCR products for chromosome Z, WL and WS make it possible to divide the intron region into 5 regions, from A to E. B1 and B2 indicate the polymorphic region. Comparison of the DNA sequences of chromosome W clones indicates that the polymorphism in chromosome W is derived from the number differences of a 32-bp-repeat. When the DNA sequences of chromosome Z and W are compared each other, the region A and E show highly homologous. On the other hands, D shows a striking difference. Region C is observed in only chromosome Z.
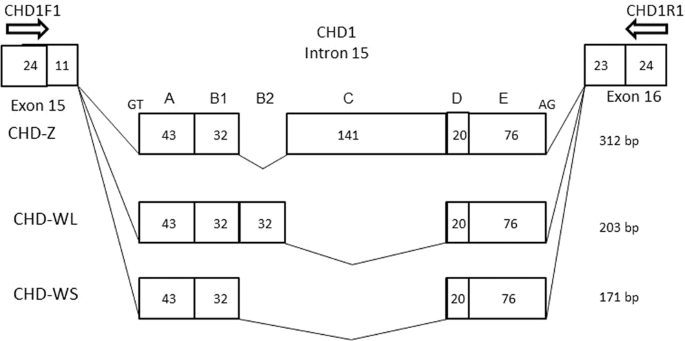
Schematic figures of the PCR products from chromosome Z and two polymorphic clones from chromosome W, WL and WS clones. The intronic regions origin at GT and end at AG as in GT-AG rule of introns. The estimated length of introns in CHD-Z, CHD-WL and CHD-WS, amplified with the primers made in this study, CHD1-F1/R1, were 312, 203 and 171 bp, respectively. Polymorphic region of the intron of GHD1 genes on chromosome Z and W of Japanese murrelet. The region containing a polymorphic intron divided into five areas, A to E. Comparison between CHD-WL and CHD-WS indicated that the DNA sequences of this region were coincided perfectly with two exception with the copy number of a-32 repeat and one nucleotide change in region A. On the other hand, comparison between CHD-Z and CHD-W, the region C exites in only CHD-Z. The regions of A and E have more than 70% homology, however, B involved repeat region and D region in a-32-bp-repeat have lower homology than regions of A and E.
The new primer set, CHD1-F1/R1, prepared in the present study is expected to be useful for molecular sexing in the Japanese murrelet, and especially using degraded DNA samples. Additionally, the results of agarose gel electrophoresis simultaneously classified the females into two groups based on the presence of a chromosome-W polymorphism. Distribution of a 32-bp-repeat polymorphism in the intron of the CHD1 gene on chromosome W of the Japanese murrelet on Biroujima Island in Miyazaki Prefecture in Japan is shown in Table 1. In the 61 females identified among 149 individuals, 9.8% of the females harbored 1 copy of the 32-bp-repeat polymorphism. The remaining 90.2% of the females harbored 2 copies of the 32-bp-repeat polymorphism.
Source: Ecology - nature.com
