A total of 318 calves were sampled across 12 farms positive for the environmental presence of VTEC O157:H7. Fifty-six calves were colonised as detected by recto-anal mucosal swabs (RAMS) from the terminal rectum and their faecal shedding levels varied between 0 and 840 000 cfu/g faeces (Table 1). Animal-based assessment of health and welfare of the 56 colonised individuals (cases) were compared to 135 non-colonised calves (controls) housed in the same pens (n = 26) to describe patterns in calf behaviour and welfare that may influence the risk of colonisation. In this way, we controlled for environmental and management factors associated with the pen. See Supplementary Table S1 for assessment-protocol.
The number of colonised animals (RAMS positive) varied between the pens (range 0–9 individuals per pen) and only 19 of the colonised animals were shedding VTEC O157:H7 in their faeces (positive faecal sample). However, most of the shedding individuals shed high levels of bacteria (68% shed >104 cfu/g, 89% shed >103 cfu/g). PCR confirmation of two individual isolates from each farm showed that all farms except one had isolates with the virulence genes eae and vtx2 (the exception had eae and vtx1). All isolates except the vtx1 positive isolate came from farms located in a region known to have a high prevalence of a virulent strain of clade 8 and were likely closely related37. The remaining isolate was from a farm that was identified in association with an epidemiological investigation following a human case. Thus, all isolates in the study have the potential to cause disease when transmitted to humans.
Visualising associations between animal-based and behavioural variables using cluster analysis
The animal-based assessments and colonisation status of the sampled animals are visualised in Fig. 1 (more detailed results are found in Supplementary Table S2). The top horizontal axis of Fig. 1 represents each individual calf (clustered by similarity, using Gowers distance) and the animal-based variables (clustered by association) are presented on the left vertical axis. Noteworthy is that the colonised animals did not form a distinct individual cluster i.e. the swab positive calves were distributed along the horizontal axis and not to specific clusters of individuals (see red box. Fig. 1). Instead, individual similarity appeared to be based on many different factors. Calves with poor coat condition appeared to cluster (see cluster a. Fig. 1) as did calves with nasal discharge (see cluster b. Fig. 1). In addition, a cluster including calves, characterised by having wounds/inflammation and poor ruminal fill was identified (see cluster c. Fig. 1). This cluster also included a high proportion of the castrated males sampled within the study.
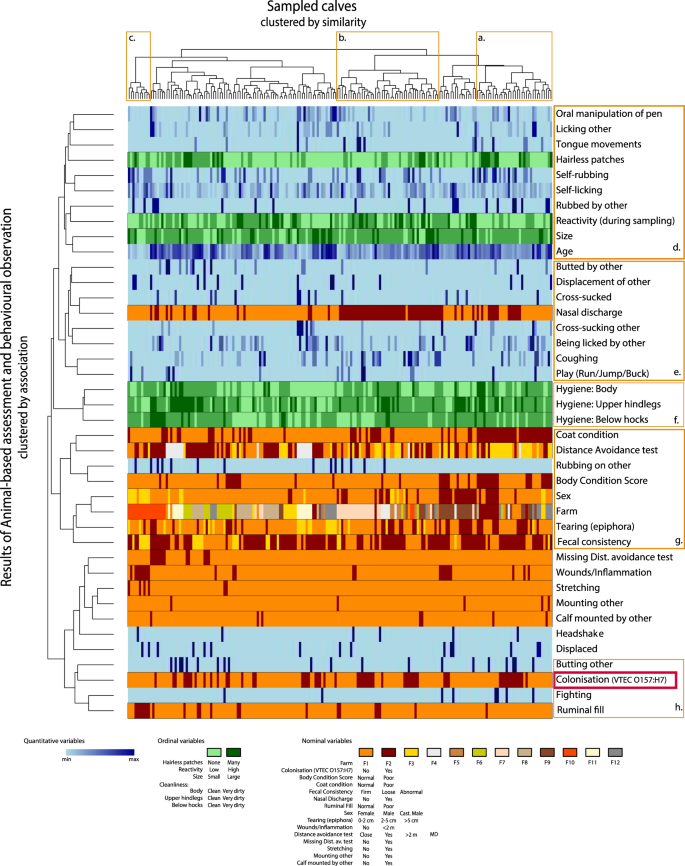
Cluster analysis including welfare and behavioural observations of the 56 colonised (positive recto-anal-mucosal swabs) and 135 non-colonised dairy calves from 12 Swedish farms infected with verotoxin-producing Escherichia coli serotype O157:H7 (VTEC O157:H7). The top horizontal axis represents each individual calf (clustered by similarity, using Gowers distance) and the animal-based variables (clustered by association) are presented on the left vertical axis. Results of the animal-based assessment of each variable (and which farm the individual belonged to) are illustrated by a vertical line below each individual. To aid references to this figure in the text, clusters are given the letters a-h.
Associations between the animal-based assessments included a clustering of comfort behaviours like self-licking and rubbing. These behaviours clustered close to variables associated with oral manipulation of peers and the environment and having hairless patches as well as age, calf size, reactivity during sampling and being rubbed by other calves (see cluster d. Fig. 1). There was also a cluster which included receiving and performing active/social behaviours like being butted and displacing others, as well as playing, nasal discharge and coughing (see cluster e. Fig. 1). Hygiene related variables; such as cleanliness of lower hind legs, hind and body, formed a distinct cluster (see cluster f. Fig. 1). Farm clustered with a number of potentially management related variables like coat condition, fearfulness as indicated by the distance avoidance test, body condition score etc. (see cluster g. Fig. 1).
Variables clustering closely to colonisation status were butting of other calves, fighting and ruminal fill (see cluster h. Figure 1). Butting others appeared to be positively associated while poor ruminal fill appeared inversely related to colonisation. This is surprising, as dietary stress has previously been correlated with increased shedding post inoculation of a large number of VTEC O157:H738. This may indicate that the effect of fasting is different for colonisation compared to shedding. The association with fighting is difficult to assess due to the low frequency of calves performing fighting behaviour (n = 8).
Identifying predictors of colonisation status using elastic net regression
Results from the behavioural and welfare assessments of the individuals, and pen-ID, were included in an elastic-net logistic regression model with colonisation (yes/no) as response variable. Coefficients for maximal area under the curve (AUC) after 10-fold cross-validation are presented in Fig. 2. Pen-ID was included to account for the fact that calves were kept in groups and for physical characteristics of pen. The identified individual variables are therefore relevant for all calves in any pen. The inclusion of pen-IDs in the final model indicates that, in addition to individual calf differences, there are pen level characteristics (social or physical) that influence colonisation risk. Understanding such characteristics may be important for reducing the prevalence of VTEC O157:H7 and should be considered in future studies.
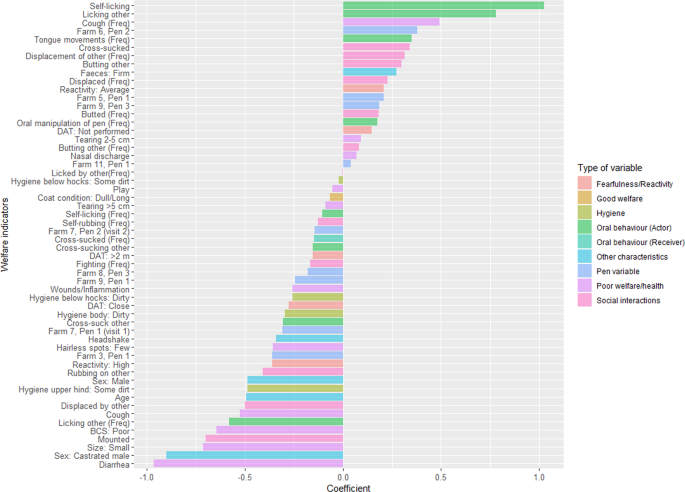
Coefficients in the elastic net regression model that maximised area under the curve (69.3%). Positive coefficients were positively associated with colonisation by verotoxin-producing Escherichia coli serotype O157:H7 and negative coefficients were negatively associated with colonisation. Behavioural variables marked with (Freq) are quantitative measures of the number of times the behaviour was observed as opposed to binary variables indicating whether a behaviour was observed (yes/no). The variables are grouped to help illustrate how the different types of variables are associated with colonisation.
The associations identified by the elastic net regression model were partly consistent with the cluster analysis. For example, butting others was positively associated with colonisation while mounting others and being displaced by others were negatively associated with colonisation. The elastic net regression also indicated positive associations between colonisation and oral behaviours, like self-licking and licking others. Performing these behaviours will of course increase exposure to the bacteria. Negative associations were observed with being small, castrated, having a poor body condition score and diarrhoea (firm faeces was positively associated with colonisation). This is consistent with a previous study observing a cohort of heifers on pasture where firm faecal consistency and high body condition score was associated with shedding of VTEC O157:H739.
To handle the high frequency of zero observations in some of the behavioural variables, binary variables describing if an individual calf performed a behaviour (yes/no) were created and analysed alongside the quantitative variable of the number of times the behaviour was observed40. There were several examples where the binary variable had an opposite relationship with the outcome than the corresponding quantitative variable. For example, being a calf observed to perform self-licking and licking other calves was positively associated with colonisation while the number of observations of licking were negatively associated with colonisation. This indicates a difference between calves performing these behaviours at lower frequency compared to calves performing them at high rate, although this needs to be confirmed since the negative coefficient for self-licking frequency was relatively small.
Similarly, coughing, being displaced and displacing others also had opposite relationship between categorical and quantitative variables. Being observed to perform these behaviours was negatively associated with colonisation, but within the group of animals, performing the behaviour more often increased the risk of colonisation. For coughing there are multiple reasons for increased frequency of coughing. The most obvious is a more severe respiratory infection leading to coughing, but during observations calves often coughed in association with playful activities. This association is also evident in the cluster analysis where coughing and play were together in the same cluster. Thus, frequent coughing may be a poor indicator of respiratory disease as the higher frequencies are very influenced by a small number of playing individuals. The association between colonisation and increased frequency of displacements could be differentiating between calves that are displaced once, and then do not engage in further interaction, and calves that are actively participating in competition and social interaction with their pen-mates. Indeed, the overall picture when looking at all coefficients, both positive and negative, indicates that the colonised calves are not found among the sick, stressed and so potentially immunocompromised calves in a group, as hypothesized, but among the animals that are active, grooming and socially engaged. This is in keeping with the previous analysis showing that colonised calves did not cluster with those showing symptoms of sickness or stress, but close to those butting other calves, fighting and having good ruminal fill.
Exploring interdependence and covariation using principal component regression
The 39 variables from the animal-based assessment were combined using principal component analysis (PCA). Thirty-two principal components (PC) with eigenvalue > 0.6 (explaining 87% of the variation in the data, each component between 5.7–1.3%) were subsequently included in a logistic regression. After backwards stepwise selection using Akaike information criterion (AIC) the final PC regression model included 14 PCs associated with colonisation of which 7 had a p-value < 0.05. Estimates from the logistic regression are presented in Table 2. The variable contributions (in percent) to the significant PCs are presented in Fig. 3. Contributions of all components included in the model can be found in Supplementary Fig. S3.
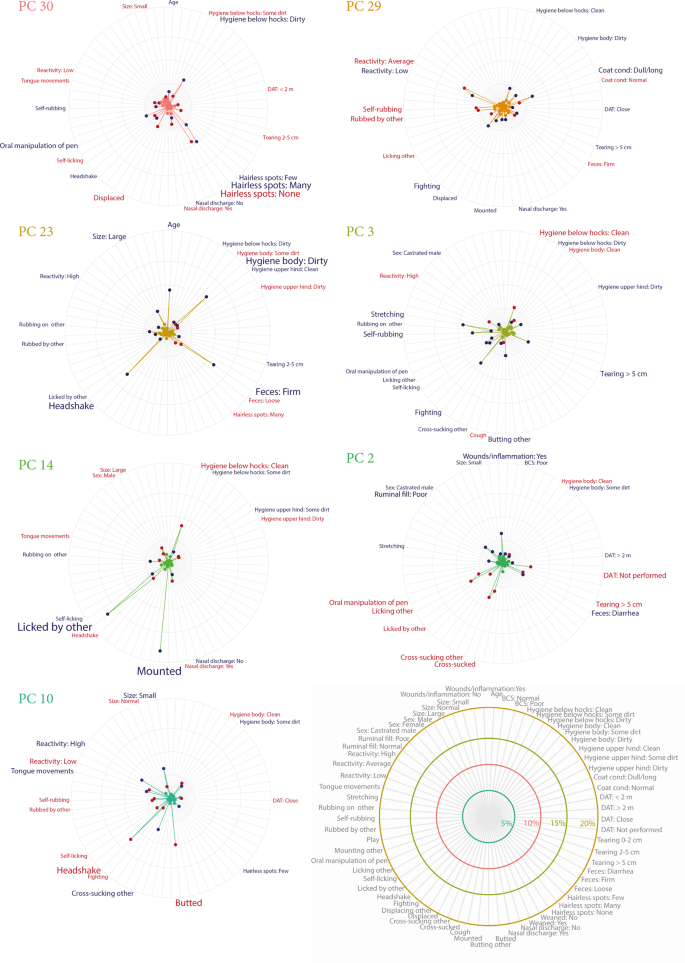
Variable contributions (%) for the seven significant principal components included in the final principal component logistic regression model. Variable contribution indicates how much influence the individual variables have within each principle component – the longer the line (i.e. the greater the %) the more influential that particular variable. Red variables are positively associated with colonisation of verotoxin-producing Escherichia coli serotype O157:H7 and blue variables negatively associated. Each principle component represents a different aspect of the variation in the whole dataset, so the variables within each component should be interpreted together. DAT = Distance avoidance test.
The importance of several associations suggested by the elastic net analysis are confirmed in the significant components of the PC regression. The association between social behaviours and poor welfare are especially clear within PC 2 (p = 0.038) (Fig. 3) where oral manipulation of the environment as well as performing and receiving grooming (licking) and cross-sucking was positively associated with colonisation. In contrast indicators of poor welfare (like wounds/inflammation and poor ruminal fill) were negatively associated with colonisation. Interestingly this PC indicates a correlation between the social and oral behaviours and that the distance avoidance test (DAT) not being performed (due to restricted space) which may indicate that these types of behaviours are occurring more frequently in smaller, more densely populated pens. Self-licking, which received the largest coefficient in the elastic net regression, appeared in several of the significant PCs but was not among the most influential contributors on any except for the non-significant PC 8 (p = 0.071) where self-licking was the second largest contributor (Table 2). This component also indicated a correlation between self-licking, rubbing, oral manipulation of pen and displacing others and that these were linked with colonisation (Supplementary Figure S3).
Other PCs that revealed a pattern similar to PC 2 (i.e. appeared to separate calves with good health and welfare from calves that were coping less well) were PC 30 (p = 0.004) and 29 (p < 0.014). PC 30 differentiated between calves without hairless spots and calves that were dirty below the hocks and had few or many hairless spots. PC 29 separated calves with a normal coat and average reactivity, performing self-rubbing, from calves with a poor coat condition and low reactivity. Looking more closely at these components, they also visualise the complex interdependencies within the data set. For example, in PC 30 being displaced and having low reactivity were positively associated with colonisation but on PC 29 they were negatively associated with colonisation. This suggests that these variables have context dependent relationships. This may relate to the finding that calves displaced more frequently were more at risk for colonisation compared to calves displaced at lower frequency observed in the elastic net regression.
Component 23 (p = 0.025) is most likely describing a protective effect of increasing age which has been previously observed41. It also highlights a correlation between increasing age, dirtiness and firm faeces. This association may explain why poor hygiene appears to be negatively associated with colonisation on multiple components. However, considering the cross-sectional design of this study it could also be that calves kept in a dirty environment, with high infection pressure, actually may have been previously colonised and therefore less likely to be colonised again due to immunity.
Being a male or a castrated male was associated with reduced risk of colonisation in the elastic net regression. While some previous studies have identified castrated males as less likely to shed VTEC O157:H742 and that heifers and cows are more likely to be colonised17, others have identified very small differences between sexes41. In the PC regression, being a castrated male was associated with reduced risk in PCs 2 and 3 and being a non-castrated male was associated with increased risk in PC14. Due to the small number of observed castrated male calves (n = 19) it is likely that the associations observed are a result of correlations between variables and thus not a direct effect of sex on colonisation risk. Similarly, PC 14 is highly influenced by being mounted by others. This variable was picked up by the elastic net regression as well as by the cluster analysis but as it was observed at very low frequency (n = 7) there is also a risk for spurious associations.
General discussion
The aim of this study was to investigate the association between welfare and colonisation of VTEC O157:H7 as well as explore patterns of behaviour and welfare to identify risk behaviours. The main hypothesis was that calves with indicators of poor welfare are more likely to be experiencing stress and stress-induced immunosuppression and therefore more susceptible to colonisation compared to other individuals in the same pen43,44. However, several variables traditionally used to define poor welfare were negatively associated with colonisation. This was especially clear in the results of the elastic net regression where, for example, having diarrhoea, being displaced and having poor body condition score were negatively associated with colonisation. This was supported by the principal component regression where similar patterns were observed within several significant components. Thus, there was no support for the suggestion that individuals with poor welfare are more likely to become colonised. In fact, the results seem to support the opposite view, that it is active, social individuals (involved in agonistic interactions and possibly high exertion play movements) who are colonised.
There were indications that colonised animals may be more frequently exposed to the bacteria by performing more comfort behaviours (i.e. self-licking) and by grooming others. Self-licking is a behaviour with many motivators45 but one of the most obvious drivers is a direct response to being dirty, meaning that the calves that are in a dirty environment and are licking to clean themselves are more likely to ingest the bacteria. These calves would be cleaner which could explain why hygiene is not emerging as a risk factor in our analysis. In addition, grooming and cleaning behaviours require an energy investment and it is for this reason that sick individuals do not groom themselves as often46. This could explain the association between non-colonised animals and poor health and welfare observed in both the elastic net regression and PC regression.
Social grooming, besides also increasing exposure to the bacteria, plays an important role for tension relief as well as maintaining and building relationships46. The observed connection between colonisation and calves performing social grooming and agnostic behaviours are interesting from a disease spreading perspective. The distinction between super-shedders (individuals shedding high levels of bacteria) and super-spreaders (individuals with more contacts/interactions with other hosts) as independent traits has previously been introduced for VTEC O157:H713. Results of this study are the first to provide evidence that risk factors for colonisation are also associated with behaviours related to social contacts and interactions with peers, i.e. risk behaviours for super-spreading.
From the PC regression, it is clear that there are complex interdependencies among the predictors that jointly influence the colonisation process. A common method for handling many variables is reducing the data by removing some of the variables based on biological reasoning and the focus of the research question. However, when prior information is lacking the decision of which variables to remove can be difficult and lead to oversimplification and bias. In addition, the possibility to consider associations between variables, which may provide important information for interpretation of results, disappears. In this study we approached this by combining multiple methods with different strengths and weaknesses. Combining the results of all methods revealed similar patterns and the methods strengthen each other as well as reveal different aspects influencing the analysis.
As a first step we used cluster analysis to visualise associations and data. This provided a good overview of variables and indications of correlation and associations. However, there was no way to assess goodness of fit and clusters may change drastically if variables are excluded. The elastic net regression enabled analysis of all variables (including the 26 pens as fixed effects to control for non-independence within pens) and provided a reduced model that still included correlated variables, like self-licking and licking others, even if this induced variance inflation when combined together in a logistic regression. However, the limitation of this approach remains that the coefficients generated are biased estimates and cannot be used to calculate odds ratios to quantify the importance of the predictors. In addition, the area under the curve (AUC) for the best model presented in this paper was 69.3%. This means that the individual variables only gave us part of the picture. The AUC for the PC regression, on the other hand, was 81.2%, indicating that the inclusion of non-independent data increased model performance and was better at capturing the complex interdependencies of welfare metrics. In addition, it exposed interesting correlations between variables, deepened the understanding of associations and revealed potential sources of bias in the study. However, the limitation of the PC regression method is that the creation of components is driven by variation within the data and therefore unrelated to outcome. There may be variables that are strong predictors for colonisation, but if they do not explain variation within the data they will not be represented on any component and so will not be identified. Another drawback of this method is that it can be difficult to infer the biological pattern represented by each components. The strength of using these different approaches is the increased confidence in associations identified within all three methodologies. Nevertheless, it should be noted that some associations may have been missed due to the relatively short behaviour observation period or the number of animals (i.e. there is a risk of type II errors).
Although the cross-sectional study design means that causality cannot be inferred we have managed to tease apart the complex structure of individual patterns. We suggest that future studies test the hypothesis that performing behaviours related to oral exposure, like self-licking and social grooming, increase the risk of colonisation in calves. In addition, the underlying association between poor welfare indicators and reduced risk of colonisation should be further investigated to identify if this is due to immunity from earlier infection (with this or similar strains of bacteria), decreased exposure (e.g. due to sickness behaviour) or differences in colonisation resistance (e.g. variations in the microbiome or other features of the terminal rectum of these individuals). As others before47, we suggest longitudinal studies with increased frequency of observations that will enable the investigation of how individual colonisation resistance/susceptibility changes over time in relation to changes in behaviour, previous infection and increasing age. The individual drivers of colonisation emerging in this study have important implications for understanding of VTEC O157:H7 transmission within farms as well as for developing on-farm measures for reducing the prevalence of the bacteria on farms in the future.
Source: Ecology - nature.com
