Experimental results
During our weathering experiment, observed for a period of 12 months, the glass lizard carcass remained in complete articulation with scales for the first two months (November and December, 2018), showing no major changes; descaling subsequently occurred but the skeleton remained in complete articulation. After three months the skeleton started to disarticulate, with some bones separated from the main body of the lizard. The most drastic change occurred after five months when most of the skeleton was disarticulated. After nine months in the field the bones were mostly disarticulated (Supplementary Fig. S1). Only very slight cracking of the ventral surface of the glass lizard vertebrae appeared after nine months, while the digested bones showed no change at all. After one year the cracking of the glass lizard vertebrae ventral surfaces progressed to Andrews’ stage 1 (Supplementary Table S1). No bone surface modifications (e.g. cracking) were observed on the digested bones at any point in the experiment. The experiment is ongoing and is now in its second year.
Our burning experiment produced cracking of bone surfaces, mainly on the condyle, cotyle, parapophysis and diapophysis of vertebrae (Supplementary Fig. S2). Greater burning intensity (time × temperature) positively correlated with more extreme cracking (Spearman’s r = 0.97 p < 0.01). Natural and digested bones were found to have been similarly affected in each time/temperature combination (Supplementary Table S2, Supplementary Fig. S3). Squamate vertebrae appear to turn black in lower temperatures than fish cranial remains30 and macro-mammal bones31,32 perhaps due to their smaller size and larger surface area. Owl digestive process does not appear to have an impact on the bone charring temperature.
Erosion by sediment produced diagnostic marks on the diapophysis, parapophysis and condyle to varying degrees. On untreated bones the slightest erosion was observed as slight abrasion of the bone surface. Moderate erosion occurred as abrasion of the bone surface that produced slight perforations with irregular edges on the protruding edges of the diapophysis, parapophysis and on the condyle. Great erosion occurred as large perforations with irregular edges on the entire surface of the diapophysis, parapophysis and on the condyle, and extreme erosion occurred as complete abrasion of the diapophysis and the parapophysis and large perforations with irregular edges on the condyle. On digested bones slight erosion was observed as slight abrasion of the digested bone surface and smoothing of the regular-edge perforation on the protruding edges of the diapophysis and parapophysis. Moderate erosion occurred as the abrasion smoothed the regular-edge perforation on the entire surface of the diapophysis, parapophysis and condyle. Great erosion occurred as parts of the diapophysis and parapophysis were broken and rounded along with the smoothing of the bone surface and, like the untreated bones, extreme erosion occurred as complete abrasion of the diapophysis and the parapophysis (Supplementary Fig. S4).
Overall, longer rotation time led to greater erosion of the bone surface (Supplementary Table 3; χ2 = 3.85, p = 0.05). Some variability was observed according to the state of the bone (untreated, digested or burnt). In the untreated sub-sample, the longer the erosion time the more abraded the bones surface became. Burnt bones were found to be more susceptible to erosion, with extreme erosion in three out of seven specimens. The digested bones produced variable erosion irrespective of time.
Trampling similarly produced abrasion of the bone surfaces and perforation with irregular edges on the diapophysis, the parapophysis and on the condyle (Supplementary Fig. S5). Trampling also produced breakage of the protruding parts of the vertebrae (diapophysis and parapophysis, prezygapophysis, prezygapophyseal articular facet, hypapophysis and postzygapophyses), which we scored from 0 (all the protruding parts were complete) to 7 (all the protruding parts were broken). Here too, the state of the bone (untreated, digested or burnt) appeared to have had an impact. Untreated and digested bones were rarely broken, while burnt bones had a noticeably lower completeness index and higher protruding-part breakage index: of 17 burnt specimens, one was not broken at all, three had a low breakage index, six had a moderate breakage index, four had a high breakage index and three were broken beyond retrieval (Supplementary Table S4, Supplementary Fig. S6). Our pilot erosion-by-sediment and trampling experiments revealed the overall impact that these processes would have had on squamate vertebrae, but further experimentation is needed to verify the extent of variability among the untreated, digested and burnt bones.
Our actualistic observations included eagle owl pellets collected from two locations in northern Israel, Mount Nitai and Nahal ‘Ayun. Indicative digestion marks were found to varying degrees in both eagle owl pellets accumulations. The marks consist of perforations of the bone surface with regular edges on the condyle, cotyle, diapophysis and parapophysis (Supplementary Fig. S7). Of the specimens in the pellets, 81% displayed these marks; of those, half were of low intensity, 22% were of moderate intensity and high intensity was very rare (1%, n = 4 of 152 in total) (Supplementary Table S5). Breakage patterns showed that the vertebrae in the pellets were mostly whole or only slightly broken (complete = 74%, slight = 25%) with one vertebra with moderate breakage intensity (1%).
In the presumed owl pellet remains retrieved from a niche in the cliff overhanging the el-Wad Terrace by L. Weissbrod33, henceforth el-Wad Niche, the previously identified indicative digestion marks (perforation of the bone surface with regular edges) were abundant (51%, n = 138 of total 273). Thirty-three percent (n = 89) were of low intensity, 12% (n = 33) were of moderate intensity and, like the eagle owl pellets, high intensity was rare and appeared only on 5% (n = 13) of the specimens (Supplementary Table S6). In addition, perforations with irregular edges, which in our experiment had resulted from erosion by sediment and trampling, were observed in varying intensity; most of the assemblage (53% n = 142 of total 268) had slight to moderate erosion intensity, great intensity was less common (11% n = 29) and extreme intensity was very rare (1% n = 2). Breakage patterns showed that the vertebrae of the assemblage were mostly whole or only slightly broken (complete = 27%, slight = 38%) but moderate (19%) great (10%) and extreme (6%) breakage intensities were also noted (Supplementary Table S7).
Typology of bone surface modifications on squamate vertebrae
Following the experiments and observations on the pellets, we identified five bone surface modification types and linked them to different accumulation and modification scenarios, as follows:
Type A. Perforation of the bone surface with regular edges on the condyle, cotyle, diapophysis and parapophysis along with smoothing and polishing of the bone surface (Fig. 2a). Similar marks were identified previously on squamate bones10 and on micro-mammals (e.g.34,35) and were attributed to digestion. Our observations confirm their assignment to digestion, as they were observed only in the pellets and in none of the other experimental sets.

Bone modification types in the experimental material (left column) and their equivalents in the archaeological assemblage (right column). (a) Regular-edge perforations from the Mount Nitai pellet; (b) irregular-edge perforations from the trampling experiment; (c) smoothing of the regular-edge perforation from the trampling experiment; (d) cracking of the bone surface from the weathering experiment; (e) breakage of protruding parts of the vertebrae from the trampling experiment; (f) linear striations from the archaeological sample; (g) flaking of the bone surface from the archaeological sample.
Type B. Perforations with irregular edges on the bone surface that were located on the condyle, diapophysis and parapophysis and rarely on the cotyle (Fig. 2b). We produced this modification type by the erosion-by-sediment and trampling experiments. Type B was not found in any of the fresh eagle owl pellets, but was found in 45% of the specimens of el-Wad Niche, mostly to a slight degree (20.6% of the total assemblage). Owls trampling in their roost and fluvial transport may be the sources of such impact in the Niche assemblage33.
Type C. Smoothing of the regular-edge perforation (Type A) (Fig. 2c). This process was observed when digested bones were subjected to erosion by sediment and trampling (see Supplementary Figs. S4, S5).
Type D. Cracking, splits and cracks that penetrate the surface of the bone but do not cause loss of surface tissue35 were observed on the bone surface on the condyle, cotyle, diapophysis and parapophysis (Fig. 2d). We observed this modification in the burning experiment. In addition, slight cracking of the ventral surface of the vertebrae occurred in the advanced stages of our weathering experiment.
Type E. Breakage of protruding parts of the vertebrae (diapophysis and parapophysis, prezygapophysis, prezygapophyseal articular facet, hypapophysis and postzygapophyses). This damage was produced in the erosion-by-sediment and trampling experiments on untreated, digested and burnt bones (Fig. 2e). Weathering, and burning per se, did not create this damage.
In addition, two modification types, which were observed on our archaeological material (see below) but not in our experimental dataset, should be mentioned:
Type F. linear striations with a v-shaped cross section on the lateral left postzygapophysis (from posterior view; Fig. 2f).
Type G. Flaking, the loss of the bone surface tissue, described for mammal bones by Fernandez-Jalvo and Andrews34 as the result of weathering and mammalian and raptor digestion) (Fig. 2g). Our experiments did not replicate this modification.
The archaeological case study
The EN of EWT yielded the NISP of 2992 squamate remains, from a sample of 9146 archaeofaunal remains (33%); the 1-mm sample added 599 NISP. The LN sample yielded the NISP of 939 squamate remains, from a sample of 2419 archaeofaunal remains (39%); the 1-mm sample added 539 NISP. The squamates of the assemblage are represented mainly by vertebrae (95% of the total assemblage), supplemented by some skull pieces. Of the total NISP, 2041 specimens were identified to the species level and the rest to family or to sub-order (Table 1). To prevent size biases only excavation units (baskets) that underwent both 5-mm and 1-mm collection were used for taxonomic evenness and vertebra size comparisons (Supplementary Table S8).
The most common identified species in the assemblage as a whole (Table 1) was the European glass lizard (Pseudopus apodus, NISP = 691, 15.9% of the total assemblage) followed by the large whip snake (Dolichophis jugularis, NISP = 596, 13.7%), the eastern Montpellier snake (Malpolon insignitus, NISP = 371, 8.5%) and to a lesser degree the common viper (Daboia palaestinae, NISP = 96, 2%). Taxonomic evenness varied between the EN domestic contexts of the site (Inside and Outside structure II and Loc. 67 area) and the non-domestic Loc. 25 (Simpson’s index of 0.71 for the domestic contexts and 0.79 for Loc. 25, Diversity Permutation Test P < 0.05) (Supplementary Fig. S4). Within the domestic contexts, Loc. 67 area has the lowest evenness values (Simpson’s index of 0.65) and has significantly lower evenness values than Loc. 25 (Diversity Permutation Test P < 0.05). The relative abundance of taxa from these two contexts also differs, with higher abundance of the glass lizard in Loc. 67 (22%) and considerably fewer glass lizards and more colubrids in Loc. 25 (Supplementary Fig. S9). There is also a clear size difference of the centrum length of the vertebrae between the various contexts at the site and the el-Wad Niche sample (one-way ANOVA F = 283 P < 0.05). The significant difference is between the larger vertebrae of the EN domestic contexts and the smaller-sized vertebrae of the non-domestic Loc. 25. The el-Wad Niche assemblage vertebrae are considerably smaller than all other assemblages (Tukey’s Q P < 0.05) (Supplementary Fig. S10).
We now turn to reconstructing the squamate accumulation processes at the site, according to our experimental results. Three of the bone surface modification types that were defined above, types A, C and F, can shed light on the agents of accumulation. Types A and C were both identified as indicating digestion. Type A is very rare at the site (n = 9) but type C is more common (16–33% of the samples; Supplementary Table 7). Digestion abundance and intensity are significantly different in the archaeological vs. pellets datasets (Supplementary Fig. S11). While our Natufian samples yielded 16–34% digested remains, the presumed barn owl pellet remains in the el-Wad Niche yielded 41% digested specimens, and the eagle owl pellets exhibited ca. 60–80% digested specimens (Supplementary Table S7). In addition, low-intensity digestion was significantly overrepresented in all pellets, and severely underrepresented in the three domestic EN samples. Interestingly, the inside context displays abundant signs of moderate and high-intensity digestion. In contrast, the outside, Loc. 67 area and the LN samples show the lowest digestion abundance (24%, 20% and 26% digested bones, respectively) and underrepresentation of all digestion intensities.
Digestion intensity also varied between the common species at the site. The European glass lizard and the large whip snake had low levels of digestion marks, while the eastern Montpellier snake and the common viper both had overrepresentation of moderate and high intensities of digestion. By comparing the digestion intensity of the common species to the actualistic references, a clear distinction was revealed between the European glass lizard and the large whip snake, and the other species (Supplementary Fig. S12).
Type F, linear marks with a v-shaped cross section on the lateral left postzygapophyses (Fig. 3) were observed on seven non-digested trunk vertebrae originating from outside structure II (n = 5) and Loc. 67 (n = 2) and identified as large whip snake (Dolichophis Jugularis, n = 6) and in one case, eastern Montpellier snake (Malpolon insignitus, n = 1). The unique linear marks that consistently appeared on the lateral left postzygapophyses were found only in the archaeological material and were not found in the pellets, el-Wad Niche or experimental assemblages (Supplementary Table S7). Linear marks with a v-shaped cross section may result of natural agencies such as trampling36, but their relative depth, infrequency and repetitive location on the vertebrae may suggest that they constitute butchery marks, previously unknown for snakes in the Levantine record. This suggestion should be further tested in the future, and, as more squamate archaeofaunas are studied taphonomically, similar modifications may be found.

Type F, linear striations on the lateral left postzygapophyses: (a) Drawing of location of the mark; (b–j) photos of the striations.
Following accumulation, the squamate assemblage at the site was exposed to several post-discard agents of modification. Erosion of the bone surfaces is common in both non-digested bones (type B) and digested ones (type C) in all the contexts of the site (Supplementary Table S7). The intensity of erosion slightly varies: the outside context showed higher levels of erosion intensity than all the other contexts (97% of the bones are eroded, 37% of the bones have moderate erosion, 41% great erosion and 8% extreme erosion) while the non-domestic Loc. 25 had slightly lower erosion intensity (90% of the bones are eroded, 42% of the bones have moderate erosion, 21% great erosion and 0.5% extreme erosion), both higher in intensity than any of the experimental assemblages. Erosion was not observed in the fresh pellets and was considerably less common in the el-Wad Niche assemblage (65% of the bones were eroded, 33% were slightly eroded, 20% were moderately eroded and 11% were greatly eroded) (Supplementary Fig. S13).
Cracking (type D) and flaking (type G) were extremely common, but in slightly varying degrees, in all the archaeological samples (>90%). Most of the bones in the el-Wad Niche had slight or no flaking or cracking of the bone surface (Supplementary Figs. S14, S15). The abundance of digestion levels in the assemblage was compared to both cracking and flaking levels and no correlation, or a very weak one, was found (digestion vs. cracking, r = 0.019, p = 0.24; digestion vs. flaking, r = 0.137, p < 0.05) suggesting that there is no clear relationship between digestion intensity and cracking and flaking intensities.
A significant difference was discerned between the breakage patterns of the archaeological material and the actualistic assemblages (the protruding parts breakage index, Kruskal-Wallis’s H = 10.98 P < 0.05). The archaeological material had a considerably higher breakage index (between 0.43–0.67) than both el-Wad Niche (0.31), owl pellets (0.03, 0.09) and the experimental material (between 0.07 and 0.28) presumably due to the observed sediment erosion and trampling processes (Supplementary Fig. S16).
To obtain a holistic picture of the variance in taphonomic signatures of the archaeological and actualistic samples, we performed a Principal Components Analysis (PCA) of the aforementioned modification types. The first PC, accounting for 49% of variance, shows that the archaeological assemblages clustered together relative to the actualistic ones, and are clearly divergent from them (Fig. 4). The second PC, accounting for 22% of variance, also separates the archaeological samples from the pellet assemblages. The Loc. 25 sample, while clearly separated from the actualistic assemblages, diverges slightly from the other archaeological assemblages suggesting a somewhat different taphonomic history.
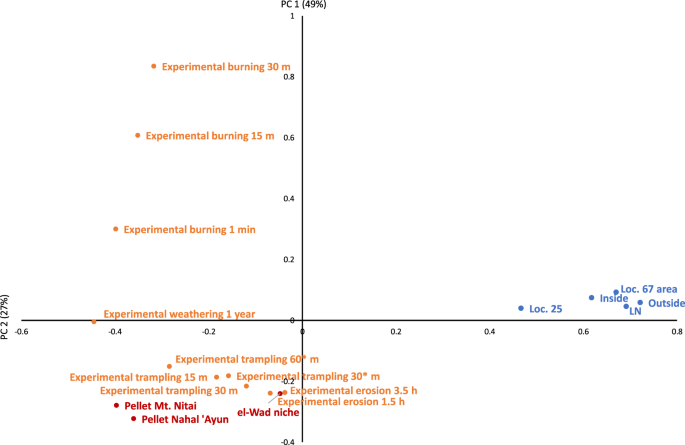
Results of Principal Components Analysis (components 1 and 2) of the different bone modification types in archaeological contexts (blue), pellet assemblages (red) and experimental material (orange).
The four common species from the entire EWT archaeological assemblage (European glass lizard, large whip snake, eastern Montpellier snake and the common viper) were also plotted (Fig. 5). The first PC, accounting for 76% of variance, separates the European glass lizard and the large whip snake from the other two species, the eastern Montpellier snake and the common viper. The latter two cluster tightly together in PC1 and PC2, suggesting a shared taphonomic history differing from the former species.
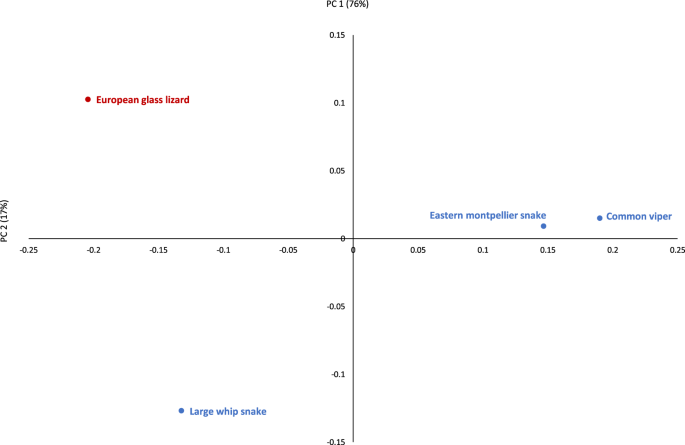
Results of Principal Components Analysis (components 1 and 2) of the different bone modification types in the European glass lizard (red) and the three other common species of the archaeological sample (blue).
Source: Ecology - nature.com
