An accurate quantification of the impacts caused by invasive organisms forms the basis for management strategies29. For fungal forest pathogens, tree mortality is the most commonly used variable to quantify losses. Since tree mortality is an inherent process of forest dynamics, defining a baseline against which mortality caused by a new pathogen can be compared presents a major challenge30. Manion and Griffin14 proposed a framework in which baseline mortality was extracted from tree inventory data collected across large areas. Because such an approach requires a lot of data, estimating mortality for minor tree species can be difficult. In this study, we developed a novel approach for minor tree species such as European ash (ca. 1% of trees in the NFI) in which mortality was explicitly modelled based on tree competition. We used pre- and post-invasion national forest inventories to identify shifts in terms of mortality and growth in ash, parallel to the spread of ash dieback. We found that in areas where ash dieback was more prevalent, plots tended to have a higher mortality than that predicted by the pre-invasion inventory – overall ash dieback increased mortality by 74% and mortality more than doubled in the smallest diameters. In areas not yet invaded, mortality mismatches amongst inventories occurred stochastically. The same pattern was found for growth, i.e. after the invasion ash had been growing 13% less than predicted by pre-invasion observations. By using the same comparative approach on other tree species co-occurring with ash in the same plots, we could exclude that mortality and growth differences in ash were due to unspecific abiotic factors, such as drought, spring frost, snow or windstorms. A higher-than-normal mortality could have been compensated by an increase of growth amongst surviving ash individuals. We observed the contrary. Higher-than-normal mortality correlated with a smaller-than-normal growth of the surviving ash trees, indicating that ash could be eventually replaced in the mid-term.
We shall also acknowledge some limitations in our approach. Firstly, the power of our analysis was limited by the low occurrence of ash trees in the plots available from the NFI. The Norwegian NFI has a grid of 3 × 3 km, and forest tree species with strong spatial structure such as ash are rarely captured as pure stands. Indeed, ash was very often recorded growing in mixed stands with other species, particularly with spruce. These conditions may not represent what happens in pure ash stands, or in mixed broadleaved stands in which disease development, and intra- and interspecific competition may be different. Coker12 estimated ash mortality rates by analysing targeted surveys reporting the proportion of trees that have died in sites across Europe and found mortality values as high as 85% in some pure European ash trials/plantations. Similarly, mortality values higher than 60% have been found in forests in Latvia and Lithuania31,32. The lower mortality recorded in the present study reflects probably country-specific differences in disease history: while the south-eastern Norwegian stands with the longest disease history were infected around 200624, the data from targeted studies with the highest mortality in Central Europe come from forest stands with a disease history of 15 to 19 years12. In addition, the level of pathogen infection pressure can be expected to correlate positively with ash stocking density and thus differ between ash dominant stands typically included in targeted studies and stands where ash is a minor species, such as most of the plots now examined. Mismatches between observations in diseased areas and NFI data have been reported also previously. For instance, work on Cryphonectria parasitica and sweet chestnut in Spain showed contrasting results when comparing high incidence in managed stands with NFI data; the results indicated that instead of declining, chestnut population was increasing in volume, likely due to management abandonment of coppice stands33.
Additionally, the records showed that some ash trees had been removed from the stands, although there was no information concerning the purpose of the removals and whether they were part of a management plan. Possible explanations were that managers opted to cut valuable timber before the disease led to deterioration of wood quality, or, alternatively, opted to remove the more damaged trees during thinning, while maintaining the healthy and potentially tolerant ash individuals34 (although, arguably, ash is not particularly managed in Norway). In the first case, this could add to the model biases, resulting in an overestimation of the mortality. In the latter, this would mean that we have underestimated ash mortality. Since there is no information about the state of the trees prior to the removals, it is not possible to effectively evaluate their consequences in the models. However, the removed trees represented a similar percentage for each diameter class and accounted for less than 1% of the total recorded ash trees, which reduces the potential bias due to the removals.
We believe that by using NFI data we can obtain a broad and largely unbiased picture that can put in perspective observations from targeted surveys. NFI are based on a standardized sampling and measurements which allow an easy comparison across large areas, compensating the effect of local conditions disturbing the analyses. Furthermore, NFI data allowed us to determine a baseline mortality for comparative purposes. Our results are consistent with observations obtained from targeted monitoring of ash dieback in Norway35; these results showed similar patterns of mortality to those obtained from NFI, with the highest mortality rate for both the smallest trees and plots in the region with the longest disease history, in the south-eastern part of the country. On the other hand, the increased number of plots with ash and ash trees between the first and the last observations, despite the mortality, deserve also attention. A potential range expansion of ash due to climate warming, land use changes or increased numbers of surviving natural regeneration2,12 may be operating in parallel to the spread of ash dieback across Norway. Further forest inventories will be needed in future to monitor how ash stands develop in the long term after the infection.
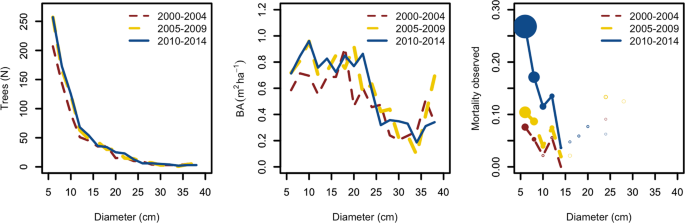
Another limitation of our data was the time lag between inventories. We found a clear trend of increasing mortality in ash, particularly reflected in the last available NFI (2010–2014). A parallel analysis of ash growth showed also a reduction of radial increment, and a weak although significant negative correlation between both mortality and growth at plot level. A plausible explanation is that ash dieback is more likely to cause tree death in smaller trees than in large trees. This has implications when fitting our models, since growth losses in smaller trees may have been masked by death, as the 5-year-period between inventories was possibly too long for those trees to survive. When comparing pre- and post-invasion inventories, growth data was not accessible for small trees because in the next inventory the trees were dead. Regional differences in tree growth may also confound those caused by a new pathogen should the pathogen spread route parallel natural gradients. In our case, the fact that ash stands in the north were possibly growing less than ash stands in the south could have affected our capacity to detect an impact. However, regional growth differences should in theory be constant across inventories, and in our case, we only observed them in the last inventory.
Baseline mortality was modelled emulating the biological process underlying tree selection in natural forests. The structure of the data allowed the use of mixed models at tree level both for the mortality and the growth analyses in pure or mixed stands. In the case of the mortality model, the variables considered and model structure was based on the model of Sterba and Monserud36 for central Europe and that of Bollandsås37 for Norway. Mortality can be modelled based on tree characteristics with or without including the context of the plot. We tried to fit both, and the predictions were more accurate when plot variables were included. The probability of dying for a tree was modelled by considering the diameter of the tree, the basal area of the plot and the underlying biological processes: (i) larger trees within a plot are the ones showing the highest vigour and a (ii) higher BA increases competition for light and water amongst trees in the plot. Without including plot variables, the models underestimated mortality, particularly for young trees.
The negative correlation between mortality and growth indicates a lack of compensation of mortality by surviving ash trees within diseased plots. Although other reasons could explain this pattern, the most plausible one is that trees in the same plot are also diseased and therefore not able to make use of the resources made available through the decline of conspecifics. This is especially worrisome considering that ash is mostly represented in mixed stands and suggests that without intraspecific resilience it may be losing its niche. Another possibility would be that mortality removed too many trees in too short a time, so that the remaining trees, even though they were liberated from competition, did not have time to respond and fill the gap caused by the disease. Further follow up of these plots should confirm whether the observed pattern persists over time or represents a transitory situation. Finally, the observed appearance of ash in new plots across the inventories demonstrates the high dispersal capacity of the tree species38.
In our study, we have shown that NFI data can be used as a baseline to quantify impacts of a novel pathogen. Although other sources of mortality cannot be ruled out, due to the limitations of NFI data, our method represents a step forward from previous approaches, as mortality is explicitly modelled on a biological basis, and demonstrates that impacts on minor tree species growing in mixtures can be quantified. Including tree growth in our analysis allowed testing the resilience of the forest to possible pathogenic invaders and to devise the consequences in the mid-term. Concerning ash in Norway, we have shown that there has been an increasing mortality and reduction in growth after ash dieback was observed in the country. Although ash is a highly prolific species with a relatively high growth rate, it may not be able to buffer ash dieback, as no sign of growth compensation by the surviving trees was observed.
Source: Ecology - nature.com
