This study is the first to provide information on the edaphic and temporal patterns associated with T. melanosporum FB traits. Our results indicated that each of the examined traits was differently affected by environmental factors. Furthermore, FB weight and Leiodes infestation were subject to interactions among these factors.
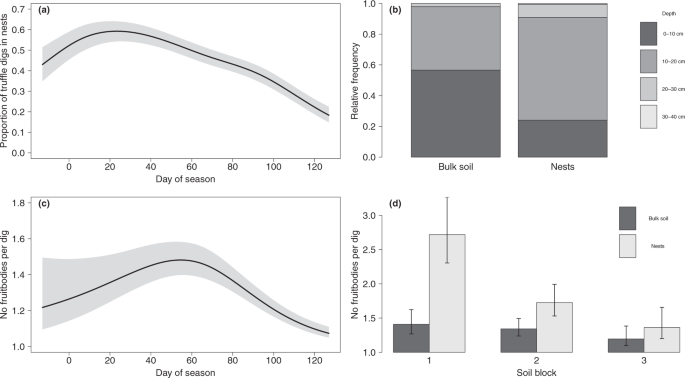
The number of FBs per dig varied over the fruiting season. If, as hypothesised by Pacioni et al. (2014)6, the optimal time for fruiting initiation is characterised by a temperature-moisture window of opportunity, our results could be related to the trends in soil conditions during spring, when fruiting initiation is typically triggered. However, our results could also be explained by time-dependent changes in FB mortality. Intriguingly, the number of FBs per dig did not show differences between seasons or among the bulk soil of the three soil blocks.
Fruitbody weight showed a more complex pattern. The weight of FBs excavated in the bulk soil in season 2016–2017 was influenced by soil properties, with more sandy soils tending to present higher weights. However, considering that in 2017–2018 no differences among soils were found, this effect appears to be strongly dependent on annual environmental conditions, either through their influence on soil microclimate or on host physiology. We hypothesise that interannual differences could be related to soil conditions or tree status during the July–October period, the FB growth stage, during which the more intense development and growth take place11,23,24. However, interannual differences in July–October temperature and rainfall in our study site were minor and restricted to the beginning of September (Supplementary Tables S9 and S10, Supplementary Fig. S2). Contrastingly, within each season no differences among the nests of the three blocks were found, suggesting that nests reduced the influence of soil properties.
Spore maturity showed a clear within-season temporal trend, with no apparent influence of the remaining predictors. According to Zarivi et al. (2015)24, spore melanisation only happens after the FB swelling is completed. Our results show a fast increase in spore maturity from early November to early December that agrees with that view, but in our case maturity continues to increase –much more modestly– throughout the fruiting season. This temporal pattern suggests that climatic conditions and/or fungal physiology rule this process, with a much lower role of soil microclimate variability. Fruitbodies fruiting in late season undergo a much longer period of low soil temperature before ripening than those fruiting in early season.
The predicted spore maturity values reached only around 80% in late season. It must be considered that gleba samples were taken from the outer part. In our experience, in early season spores and glebae are frequently darker in the inner part. Thus, our data could be underestimating spore maturity or overestimating the difference between early and late season. However, this is not likely to affect our conclusions on the remaining predictors.
Fruitbody infestation by Leiodes showed a common temporal pattern in most soil-season combinations, with maximum infestation levels in early season (although these peaks were much less marked in season 2016–2017 than in 2017–2018). This pattern could be related to Leiodes life cycle, climatic cues or abundance of FBs emitting suitable volatile signals. In a Spanish orchard, three peaks of adult Leiodes emergence were found in early, mid and late-season15. Our infestation data showed only one early peak, suggesting a relevant role of factors other than abundance of adults.
Beyond this common pattern, Leiodes infestation trend showed distinctive features in each season, with more pronounced within-season variation in 2017–2018. In both seasons, the September-February period was relatively dry, with the exception of the very rainy November 2016 (Supplementary Table S10). According to Pérez-Andueza (2015)15, adult emergence increases after rainfall, with high soil moisture enhancing the mobility of the insect in the soil. Arzone (1970)16 also pointed out that late autumn temperature drop, when accompanied by rainfall, boosts pupation. Our results also revealed that soil properties modulated Leiodes infestation trends, with the more clayey soil showing smaller amplitude of within-season variation. Clayey soils generally show lower thermal conductivity20 and soil temperature could play a role in adult emergence. Variation among soils and years indicates that soil microenvironment effectively influenced FB infestation by Leiodes.
Nests are a common practice in Spanish truffle orchards. Our results demonstrate, for the first time, that nests effectively modify several agronomically important FB traits. Fruitbodies grown in nests showed more spherical shapes and lower probability of infestation by Leiodes, thus resulting in improved truffle quality. Shape improvement could be easily explained by the lower and more uniform resistance to penetration of substrate. Lower levels of pest infestation could be related to the light, loose-structured substrate hindering the mobility of adults or impairing the bonding of eggs to soil aggregates16. This result concurs with our incidental observation, during the field sampling, that in FBs growing partly in bulk soil and partly in a nest, Leiodes galleries were mostly located on the soil side.
Nests clearly increased FB depth. This is very appreciated by Spanish growers, who feel that these FBs are less exposed to abiotic and biotic damages and are more unlikely to suffer from irregular or imperfect ripening. We have not evaluated the incidence of freezing or dryness, but in Australian orchards truffle rot is a particularly alarming problem for shallow FBs25.
Nests increased the number of FBs per dig in two of the soil blocks, with the magnitude of this effect being strongly dependent on the soil block. Murat et al. (2016)26 hypothesised that this increment could be related to spore abundance in the nest if spores were to act as male elements in sexual mating. In our study, spore abundance could be (at least partly) responsible for the differences between bulk soil and nests. However, this factor could hardly explain the different magnitude of the nest effect on each soil block, because the substrate – grinded FBs mix is prepared consistently and systematically for the entire orchard.
Alternatively, the increased number of FBs in nests could be due to enhanced FB survival. Olivier et al. (1996)27 reported that in June more than ten tiny primordia per square metre of bulk soil can be found, but during the fruiting season harvesters rarely find such clusters of ripe truffles. Considering peat properties and environmental requirements for FB growth, high water availability and aeration in nests could be pivotal in this process.
The higher number of FBs per dig found in nests could also be related to enhanced fruiting initiation. Nest installation creates an abrupt discontinuity in the bulk soil/substrate interface. Pacioni et al. (2014)6 hypothesised that fruiting initiation is triggered by a change in soil environment, which has already been found for other fungi9. In our study, the different magnitude of the nest effect on each soil supports this hypothesis. Moskal et al. (2001)28 reviewed the effect of peat amendments to soil and concluded that the increase in available water was more marked for sandy than clayey soils.
The question of whether nests increase FB weight remains unclear. Only in one soil-season combination nests affected (positively) FB weight, suggesting complex environmental interactions. This could be related to nests microclimate not always effectively improving FB growth or to a host-mediated effect.
Maturity of single-fruitbody digs was not altered by nests. This indicates that the reported issues with maturity and ripeness in nests, if true, are restricted to multiple-fruitbody digs (digs in which more than one FB was found). When a dog marks a multiple-fruitbody dig, it is only indicating that at least one FB is ripe. Considering that nests increase the number of FBs per dig in some soils, it would be interesting to investigate the presence of unripe truffles in multiple-fruitbody digs.
Fruitbody density showed in early November higher variability in nests than in the bulk soil. October is much warmer than the November-March period (Supplementary Tables S9–S11, Supplementary Fig. S2). This variability could therefore be related to water content of FBs and to the rewetting problem of peat increasing the risk of FB desiccation. Density would thus reflect the microclimate that FBs experience during their last weeks before ripening, highlighting the relevance of irrigation in peat-amended soils. This is supported by the fact that throughout season 2016–2017 density fluctuated in nests while not in the bulk soil. It would merit testing other substrates or soil mixtures that could reduce these issues.
The ratio of digs excavated in nests and bulk soil changed over the fruiting season, indicating a more heavy-tailed distribution over time in nests. Nests seem to particularly promote fruiting in early season. This could be due to the fungus finding conditions encouraging early fruiting initiation, early ripening or increasing the growth rate. Montant and Kulifaj (1990)7 found that within irrigated greenhouses (with increased soil temperature in winter-spring and increased soil moisture in summer) much more FBs were harvested in early season, with the typical soil surface cracking linked to accelerated growth of shallow FBs appearing one month earlier23. The low thermal conductivity of peat apparently points to a slow increase of temperature in spring and a slow decrease in late autumn. However, soil thermal conductivity is strongly dependent on moisture20 and peat shows high water availability at low water potentials.
The changing ratio of digs in nests and bulk soil over the season is likely to result in indirect negative effects of nests on the overall quality of truffles from an orchard, due to the time trends in spore maturity and Leiodes infestation. On the positive side, nests promoting fruiting in early December could enhance the profitability of the orchard, due to the peak of black truffle price in the weeks before Christmas.
In conclusion, our results indicate that the edaphic and temporal patterns that drive T. melanosporum FB traits are specific for each trait. The number of FBs per dig and the spore maturity showed clear within-season temporal patterns that were consistent from year to year. Contrastingly, FB weight and Leiodes infestation were subject to complex interactions among edaphic and temporal variables. In this context, nests effectively increased FB depth, improved shape and decreased Leiodes infestation, without decreasing FB maturity in single-fruitbody digs. Our results also gave hints of the issues surrounding peat hydraulic properties. The different effectiveness of nests in increasing the number of FBs among the three soils suggests a relevant role of soil discontinuities in FB initiation. The study shows that nests are a useful tool for advancing in the study of the reproductive biology and ecology of truffles.
Source: Ecology - nature.com
