Estimation of spawning time
In this study, we used two types of environmental water temperature parameters experienced by the eggs to estimate spawning time, namely the incubation temperature and the experienced temperature (Fig. 1). Arbitrary spawning time (AST) was assumed at any time before the new moon, and the time elapsed from fertilisation (ET, unit: hour) until egg collection (EC, Supplementary Fig. S1) was calculated for each egg (Eq. 1).
$$ET=EC-AST$$
(1)
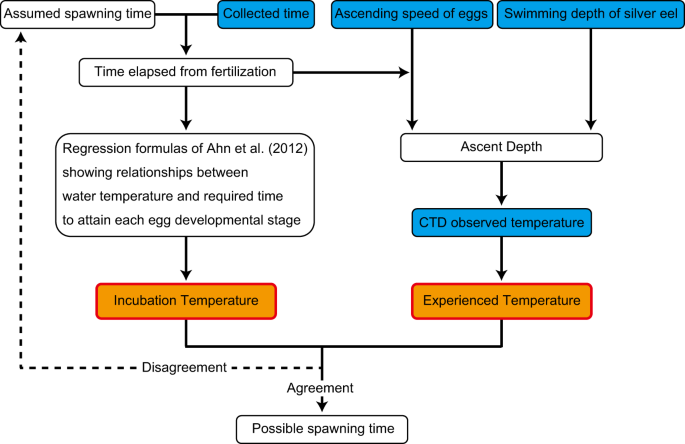
Method used to estimate the spawning time of Japanese eels. Egg collection times (Supplementary Fig. S4)10,12, ascending speed of eggs (Fig. 2), swimming depth of silver eels (Supplementary Fig. S2)18, and Conductivity Temperature Depth profiler observed water temperatures are indicated by blue boxes. The assumed spawning time (overlap of the incubation and experienced temperatures, indicated by orange boxes) is considered to be the possible spawning time.
For example, if the arbitrary spawning time is assumed at 21:00 on 3 days prior to the new moon for one fertilized egg that is collected at 3:00 on 1 day prior to the new moon, the time elapsed from fertilisation is calculated to be 30 hours.
The experienced temperature (unit: °C) is the temperature experienced by an egg prior to collection. This temperature is defined as the mean water temperature experienced by each egg during its ascent from the depth at which spawning occurred to the depth at which the egg ascent. Based on spawning depth (SD, unit: m), ascending speed (AS, unit: m h−1) and ET, the ascent depth (ADET, unit: m) was calculated (Eq. 2).
$$A{D}_{ET}=SD-AStimes ET$$
(2)
The rising arrival depth is deeper as the elapsed time from fertilisation is shorter. If the eggs reached the upper border of the pycnocline near the 150 m layer in the spawning area12, it was assumed that the eggs would stop rising at 150 m in the water column. The experienced temperature is the averaged value of the water temperature that eggs experienced during the ascending process (Eq. 3). The water temperature is the observed water temperature (Tempn, unit: °C) at corresponding depth at which the egg was present throughout the ascending process (ADn, unit: m).
$$Experienced,temperature=frac{1}{ET}mathop{sum }limits_{n=1}^{ET}(Tem{p}_{A{D}_{n}})$$
(3)
The calculated experienced temperature is lower as the elapsed time from fertilisation is shorter.
The incubation temperature (unit: °C) is defined as the water temperature at which the collected egg reached a certain developmental stage. On the basis of the relational equations between experienced temperature (variable x) and time required for an artificially fertilised egg to reach a certain developmental stage (variable y), as reported by Ahn et al.17, incubation temperature values were obtained by assigning ET to the variable y in the relational equation17. For eggs collected at different stages, the relational formulas reported by Ahn et al.17 corresponding to each developmental stage were used. (Eq. 4 for eggs at blastula stage, Eq. 5 for eggs at gastrula stage, Eq. 6 for eggs at eye and ear vesicle formation stage, Eq. 7 for eggs at heart formation stage, Eq. 8 for eggs at pre-hatching stage)
$$Incubation,temperature,(blastula)=frac{ET-11.39}{-0.2233}$$
(4)
$$Incubation,temperature,(gastrula)=frac{ET-15.383}{-0.3}$$
(5)
$$Incubation,temperature,(eye,and,ear,vesicle,formation)=frac{ET-67.822}{-1.9889}$$
(6)
$$Incubation,temperature,(heart,formation)=frac{ET-2.5222}{-88.689}$$
(7)
$$Incubation,temperature,(pre,hatching)=frac{ET-102.52}{-2.8556}$$
(8)
Thus, eggs at different stages collected at the same sampling station had different incubation temperatures. Because the egg collection time is fixed, the time elapsed from fertilisation until egg collection is variable by assumed spawning time. So, the elapsed time from fertilisation is shorter as the assumed spawning time is later, and the calculated incubation temperature is higher as the elapsed time from fertilisation is shorter.
If the assumed spawning time was the actual spawning time, the incubation and experienced temperatures were expected to coincide. Therefore, the overlap between these two estimated temperatures is herein considered to indicate the time of spawning.
The ascending speed of eggs
By observing artificially spawned eggs and their resultant preleptocephali in a cylindrical column tank, we determined the ascending speeds of eggs at the morula stage, ear and eye vesicle formation stage, and immediately prior to hatching, and of newly hatched larvae as 4.32 ± 0.11 (4.2–4.5), 3.69 ± 0.31 (3.1–4.1), 0.49 ± 0.72 (−0.6–2.0), and 7.24 ± 0.32 m h−1 (6.9–8.1 m h−1), respectively (Fig. 2). Statistical analyses revealed that the speeds of ascent differed significantly among the different developmental stages (p < 0.001, Kruskal–Wallis with Steel–Dwass post hoc test), indicating that the embryonic buoyancy decreases gradually with development and then increases rapidly after hatching18.
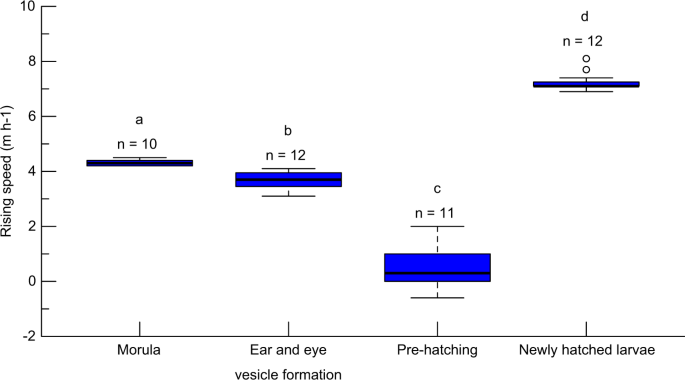
Box plots showing a comparison of ascending speeds among eggs at the morula, ear and eye vesicle formation, and immediately pre-hatching stages, and of newly hatched larvae of Japanese eels. The numbers above each box plot indicate the sample sizes for each developmental stage. Different letters above the boxes indicate a significant difference.
Eggs at the morula stage, shortly after fertilisation, may experience a temperature of under 20 °C at the estimated spawning depth (230 m) in their spawning area12, which was tracked using pop-up satellite archival tags19 (Supplementary Fig. S2). It is also known that eggs and newly hatched larvae accumulate at the thermocline (at a depth of approximately 150 m), at which the water temperature is approximately 26 °C, since the upper layer is warmer and lighter, and the eggs and larvae are unable to rise further in the water column10,12. Thus, eggs and early larvae may experience water temperatures of at least 20–26 °C. Most of the eggs collected during this study were at the late stage of development immediately prior to hatching, the measured ascending speed of which was between −0.6 and 4.5 m h−1. We used the value of an intermediate stage (3.69 m h−1; ear and eye vesicle formation stage) as the representative ascending speed of eggs in the ocean to estimate the ascent depth of the fertilised eggs.
Spawning date
The developmental stages of only 53 of the 593 eggs collected during the four R/V Hakuho Maru cruises from 2009 to 2012 were determined, as the remaining 540 eggs were either non-viable or damaged, or were unclearly photographed (photographs were taken only after a rapid onboard morphological identification) (Supplementary Fig. S3). Among the remaining 53 eggs, 7, 9, and 37 were at the ear and eye vesicle formation, heart formation stage, and immediately pre-hatching stages, respectively.
In May 2009, we collected three eggs at the ear and eye vesicle formation stage on days 1 and 2 before the new moon, and 16 eggs at the immediately pre-hatching stage on day 2 before the new moon (Supplementary Fig. S4). Using the formula described by Ahn et al.17, their spawning dates were estimated to be day 3 before the new moon (21st May 2009) (Fig. 3). Similarly, in June 2011, 2, 7, and 10 eggs at the ear and eye vesicle formation, heart formation, and immediately pre-hatching stages, respectively, were collected on day 1 before the new moon (Supplementary Fig. S4), all of which were estimated to have been spawned on day 3 before the new moon (27th May 2011). On 29th June 2011, eggs at the heart formation stage were collected at 12°54′N, 141°55′E at 04:29 h (Supplementary Fig. S3), and dead eggs were collected at 13°5′N 142°5′E at 21:32 h (Supplementary Fig. S3). The dead eggs were undeveloped although not putrid, and were thus assumed to have been spawned in the same event as the eggs collected at the heart formation stage.
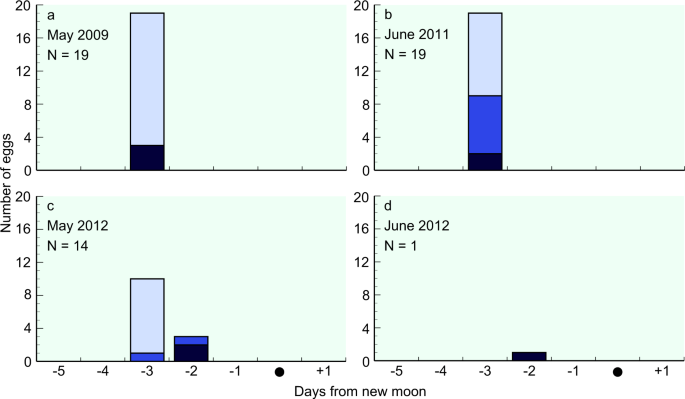
Estimated spawning date in relation to the new moon during each cruise period. The determined developmental stages of egg are as follows: ear and eye vesicle formation stage (dark blue bars), heart formation stage (blue bars), and immediately pre-hatching (light blue bars). All eggs collected are plotted in relation to the date of the new moon of each month: (a) May 2009, (b) June 2011, (c) May 2012, and (d) June 2012.
In May 2012, we collected 1, 2, and 11, eggs at the ear and eye vesicle formation, heart formation, and immediately pre-hatching stages. Their spawning dates were estimated to be on day 3 (17th May 2012) or day 2 (18th May 2012) before the new moon. One egg at the heart formation stage and 11 eggs at the immediately pre-hatching stage were collected at 15°5′N 142°20′E in the morning (07:46 h) on 19th May 2012, and on 20th May 2012, one egg at the ear and eye vesicle formation stage and one egg at the heart formation stage were collected at night (01:43–04:30 h) at almost the same location (15°5′N, 142°20′E and 15°N, 142°20′E, respectively). Given that the older hatching eggs (Supplementary Fig. S3) were collected approximately 26 h prior to collection of the younger eggs at the ear and eye vesicle formation stage (Supplementary Fig. S3) at virtually identical sites, we can assume that multiple spawning events occurred on consecutive nights within the same area. In June 2012, one egg at the ear and eye vesicle formation stage was estimated to have been spawned on day 2 before the new moon (17th June 2012). Accordingly, on the basis of the distribution of the estimated spawning dates, we can assume that spawning (94.5%) occurred on day 3 before the new moon in each spawning month.
Spawning time
A comparison between the incubation and experienced temperatures for each assumed spawning time indicated that there was no significant difference between these temperatures only between 20:20 and 22:30 on day 3 before the new moon, when the mean incubation and experienced temperatures for n = 53 eggs were 22.07 ± 3.94 °C and 23.21 ± 0.95 °C, respectively (Brunner–Munzel test, p > 0.05) (Fig. 4). Further, incubation and experienced temperatures of 4 of 53 eggs overlapped on day 2 before the new moon (Fig. 3). On the basis of this observation, we thus predict that the spawning time of the Japanese eel coincides with this time window. The 53 eggs that were used for estimation of spawning time were collected from 21:00 h to 11:00 h on days 3 to 1 before the new moon. Most of the eggs (69.81%) were collected just after the peak of spawning time (20:20–22:30 h on day 3 before the new moon) (Supplementary Fig. S1).
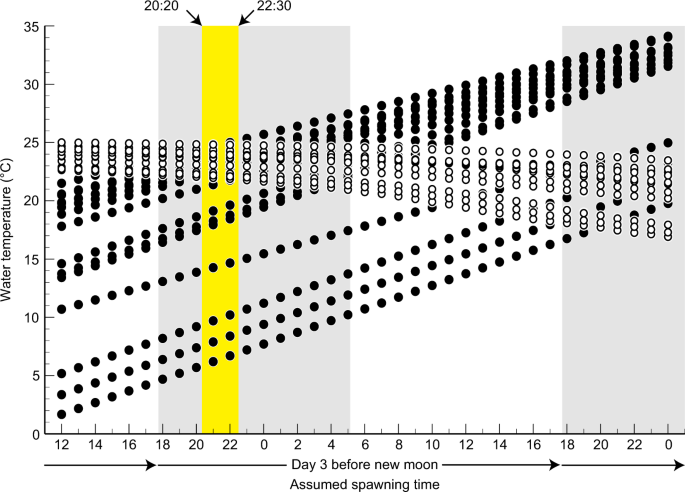
Estimated spawning time on days 2 to 4 before the new moon. The experienced temperature calculated by Eq. 3 (white circles) and incubation temperature calculated by Eqs. 4–8 (black circles) for 53 of all 593 eggs at each assumed spawning time (horizontal axis), estimated following the method shown in Fig. 1. Comparison of the two types of water temperatures that are plotted above each assumed spawning time showed no significant difference from 20:20 to 22:30 h on day 3 before the new moon (p > 0.05, Brunner–Munzel test) (yellow area). The grey-shaded areas show the time between sunset and sunrise, which is averaged among the four research cruises.
Source: Ecology - nature.com
