Influence of slope exposure on decay rates of the P. abies wood blocks
Two years after the placement of the wood blocks onto the soil mesocosms (Figs. 1, S1), the decomposition of the P. abies wood blocks was still in its early stages. This is in line with the lack of differences in mass loss within the 2-year period (Table 1). However, a lower cellulose content was reported after 104 weeks compared to the earlier time points, and such differences over time were more pronounced at the south-facing slope (Table 1). The lowest moisture content and pH values were observed at the end of the trial (Table 1). While pH was not influenced by slope exposure (Table 1), moisture was lower in the south- than in the north-exposed wood blocks (Table 1). In a representative year (2013/14), snow covered the experimental mesocosms at both slopes as shown in the temperature profile (Fig. S2). However, the mesocosms at the south-facing slope were free of snow at the end of April, while the snow cover remained at the north-facing slope until the end of May. The prolonged snow coverage and consequently the longer period at zero degrees might have affected the wood decomposition rates and the associated microbial activity at the two slope exposures.

Overview of the (A) destructive sampling procedure of the P. abies experimental wood blocks and the underlying soil (0–5 cm); and (B) wood blocks at the different time points (12, 25, 52 and 104 weeks) over the 2-year observational period at the north- and the south-facing site.
WIF richness and diversity were neither influenced by slope exposure nor time
Illumina analysis yielded a total of > 5.3 million reads (wood and soil), from which 93,546 ± 29,127 quality filtered sequences were obtained per sample after quality trimming and removal of non-target and chimeric sequences. These reads were clustered into 5,009 fungal OTUs at 97% sequence identity. The removal of rare OTUs (less than 5 reads per sample and observed in less than 3 samples) did not cause any effect on the fungal community composition (Procrustes m2 = 0.000222). The remaining 3,555 abundant fungal OTUs were retained for further analysis. The sample-based rarefaction curves indicated saturation of fungal diversity at the analysed sequencing depth for wood and soil samples (Fig. S3).
Soil samples were more diverse and had a higher OTU richness compared to the P. abies wood blocks (Fig. 2). The highest WIF richness and Shannon diversity was reported for t0 (Fig. 2). Both measures, richness and Shannon diversity, were independent of slope exposure (prichness = 0.84, pShannon diversity = 0.11) and time (prichness > 0.24, pShannon diversity = 0.24). However, under the same experimental conditions, the south-exposed P. abies wood blocks were characterized by higher bacterial richness and Shannon diversity with progressing decay28. This might be related to the faster rates of turnover in bacterial communities29, which make them more sensitive and consequently react more quickly than fungi to the physical and chemical changes as woody material decomposes. Indeed, neither WIF richness nor diversity were significantly influenced by pH (prichness = 0.42, pShannon diversity = 0.88), moisture (prichness = 0.23, pShannon diversity = 0.25) and cellulose content (prichness = 0.90, pShannon diversity = 0.86) in the two study sites. This is in line with litter transplantation experiments29, in which fungi were found to be more resistant to change and less reflective of their new environment than bacteria.
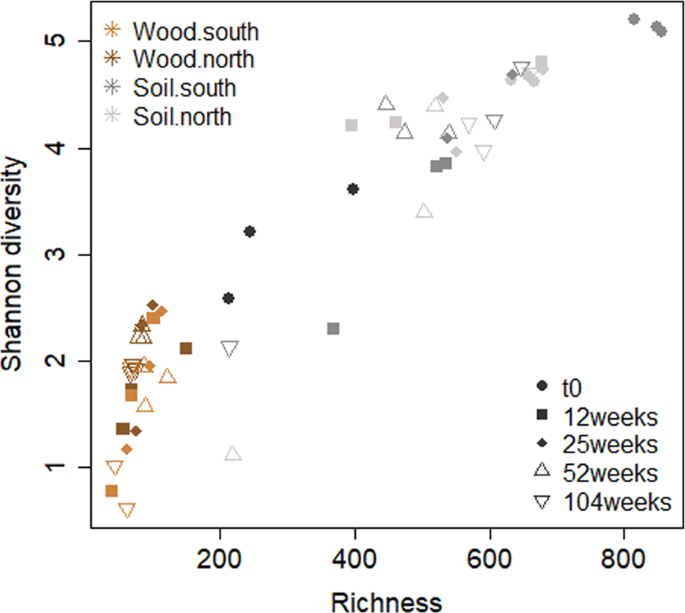
Relationship between fungal richness and Shannon diversity of the P. abies experimental wood blocks and the underlying soil samples (0–5 cm) at the different time points (12, 25, 52 and 104 weeks) over the 2-year observational period at the north- and the south-facing site.
The composition of WIF communities significantly differed from the respective underlying soil fungal communities (R2 = 0.24, pAdonis = 0.001; Fig. 3B). While the main part of the variance among the soil samples was attributed to the slope exposure (R2 = 0.24, p = 0.001; Fig. 3B), the P. abies wood blocks’ fungal composition mainly followed a time trajectory at both slope exposures (R2 = 0.33, pAdonis = 0.001; Fig. 3B). Further supporting the dependency between WIF community and deadwood decomposition progression, the experimental wood blocks’ fungal communities at the two study sites diverged over time, despite coming from the same tree (interaction effect of time and exposure, R2 = 0.11, pAdonis = 0.002) (Fig. 3B). A clearer differentiation was found between the north- and the south-exposed wood blocks after 25 (t2) and 52 (t3) weeks (ptime = 0.002, pexposure = 0.161). After 52 (t3) and 104 (t4) weeks, differences between WIF communities were also significant and accounted for over 20% of the variance (ptime = pexposure = ptime*exposure = 0.001; Fig. 3B). As occurred with WIF diversity and richness, neither cellulose nor moisture contents were significantly related to WIF composition based on the Adonis analysis (pcellulose = 0.148, pmoisture = 0.062). However, pH appeared to be correlated with changes in WIF community composition accounting for 14% of the variance (pAdonis = 0.002). Accordingly, Purahong et al.6 found wood pH as the only factor that consistently corresponded to WIF community composition in broadleaved and coniferous tree species after a 3-year decomposition time.
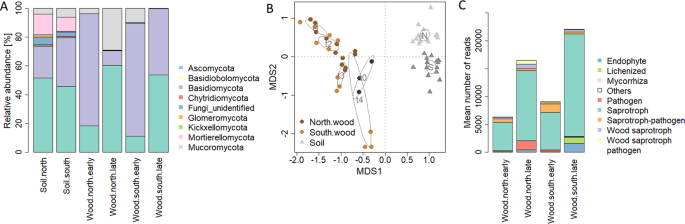
Taxonomic and functional composition of the P. abies experimental wood blocks and the underlying soil samples. (A) Distribution of taxa across wood blocks and soil samples. Operational taxonomic units (OTUs) detected by Illumina Miseq sequencing of the fungal ITS2 region were summarized based on their phylum annotation. (B) Non-metric multidimensional scaling (NMDS) based on Bray Curtis dissimilarities between the OTU compositions of the samples. Lowest stress was 0.139. The iteration converged after 20 tries. (C) The guild annotation of OTUs detected in all samples was predicted by their taxonomic annotation using FUNGuild32. The functional composition of the samples was illustrated as the abundance of OTUs assigned to the guilds. FUNGuild annotations were manually summarized into fewer categories in order to simplify visualization and data accession (Table S1).
WIF composition and indicator species differed in their taxonomy and ecologic function between the north- and the south-facing slope
In order to understand how WIF contributed to the differential deadwood decomposition, we compared the distribution of WIF taxa and their annotated functions between the north- and the south-facing site. Overall, the fungal community of the P. abies wood blocks mainly consisted of Basidiomycota and Ascomycota (36% and 53% of the sequences, respectively; Fig. 3A). Supporting Purahong et al.13, our P. abies blocks were characterized by lower numbers of Basidiomycota OTUs (meanAscomycota = 63, meanBasidiomycota = 30, pWilcox = 0.003). This is consistent with the findings by Longa et al.7, who isolated more Asco- than Basidiomycota from the experimental wood blocks analysed in the present study. In our study, the most abundant Basidiomycota OTUs belonged to Guehomyces and Mycena (54% and 12% of Basidiomycota reads, respectively). In the case of Ascomycetes, Pleosporales and Helotiales were the most abundant, accounting for 35% and 21% of all Ascomycota reads, respectively. At the earlier time points (12 and 25 weeks), the relative abundance of Basidiomycota reads exceeded those from Ascomycota at both study sites (Fig. 3A); while the opposite trend was observed after 52 and 104 weeks (Fig. 3A). In line with their abundance, the richness of Basidiomycota also decreased over time, and mainly at the north-facing slope (Spearman’s rhoNorth = −0.85, p = 5e-4; Spearman’s rhoSouth = −0.76, p = 0.004). However, the OTU richness of Ascomycota did not correlate with time (Spearman’s rho = 0.2, p = 0.33), suggesting that higher species richness is not necessarily related to faster wood degradation rates as pointed out by Rinne-Garmston et al.30.
Mucoromycota also accounted for a significant percentage of reads in the wood blocks (11%) and were almost exclusively annotated to the genus Mucor (99%). This phylum was detected on the north- and the south-exposed wood blocks at the earlier time points (Fig. 3A). However, after 52 and 104 weeks, their presence was almost negligible on the south-exposed wood blocks while it accounted for over 20% of all reads in those collected at the north-facing site (Fig. 3A). This is in agreement with the preference of Mucor spp. for colder environments31.
From a functional perspective, a total of 1,333 OTUs (956 OTUs from the filtered OTU table, that is 27% of all OTUs detected) were assigned to an ecological role using FUNGuild32. Saprotrophic fungi (general, wood and potentially pathogenic saprotrophs) accounted for 82% of reads and thus, for the majority of the functionally annotated OTUs on both north- and south-exposed wood blocks regardless of the time point (Figs. 3C; S4A,B; Table S1). However, at the later sampling points (52 and 104 weeks), WIF functional composition varied with exposure (Fig. S4C), and the south-exposed wood blocks were characterized by a higher abundance and frequency of OTUs annotated as lichenized (meannorth.late = absent, meansouth.late = 1.1%), mycorrhizal (meannorth.late = 0.03%, meansouth.late = 0.2%) and endophytic (meannorth.late = 3.8%; meansouth.late = 5.3%), and by a lower abundance of OTUs classified as potentially pathogenic (meannorth.late = 9.3%; meansouth.late = 7.8%; Fig. 3C, Table S1).
Having found differences in both WIF taxonomic (at phylum level) and functional composition between the north- and the south-exposed wood blocks, we sought to predict the indicator species that might act as key players during the 2-year period by using the linear discriminant analysis (LDA) effect size (LEfSe) tool. Considering only OTUs that were observed three times between 12 and 104 weeks, primary colonising OTUs were defined as OTUs that were present at t1 (12 weeks) and absent at t4 (104 weeks); and secondary colonising OTUs as those absent at t1 (12 weeks) and present at t4 (104 weeks). Indicator species and primary/secondary colonizers often coincided (Table S2). Among the early wood colonizers (Table S2), we mainly found basidiomycete yeasts from the classes Tremellomycetes (Cryptococcus sp., Cystofilobasidium sp.) and Microbotryomycetes (Rhodosporidium kratochvilovae). Another key player at the earlier stages was Guehomyces pullulans (= Aureobasidium pullulans), which is an ubiquitous yeast-like fungus that can colonize weathered wood, utilising products of biotic and abiotic lignocellulose degradation33. G. pullulans is very efficient in conquering the habitat in the very early stages of wood decomposition. As decomposition proceeds, its biomass can be used by other species providing a valuable nutrient source33. However, it should be noted that our experimental design focused on the decay dynamics of relatively small and equally sized wood blocks, and key attributes determining the WIF community composition include also the wood diameter34. Altogether, these circumstances could have favoured the presence and higher contribution of basidiomycete yeasts as important players during the early stages of decomposition of our P. abies wood blocks within the 2-year period.
Fungal indicator species of the later stages of wood decomposition (52 and 104 weeks), as detected by LefSe, clearly differed as a function of slope exposure. Indicator species of the north-exposed wood blocks were often Mucoromycota, as represented by five Mucor species (Table S2). Although Mucoromycota are not considered typical deadwood composers, recent research suggests that they are involved in wood decomposition via facilitating the breakdown of complex sugars35. Another interesting indicator species for the later stages of decomposition at the northern slope was the ascomycete Trichoderma viride (Table S2). Trichoderma species are poor lignin- but effective cellulose decomposers and benefit from the delignification process performed by white-rot fungi. They also act as antagonists to other fungal species, including mycoparasitic interaction in the wood using other mycelia as resources36.
At the south-facing slope, indicator species for the later stages included key players with different functions, such as the wood decomposing species Heterochaetella brachyspora, Sarea difformis and Mycena flavoalba35,37 (Tables S1 and S2). In addition, the mycorrhizal fungi Meliniomyces bicolor and Cladophialophora appeared as indicator species in the later south-exposed wood blocks (Tables S1 and S2), likely due to the earlier snowmelt and longer vegetation periods at this slope exposure. Accordingly, the underlying soil was also characterized by a higher abundance of the ericoid mycorrhizal genus Oidiodendron and the arbuscular mycorrhizal genus Ambispora (Tables S1 and S2).
Associations of N2-fixing bacteria and fungi were more frequent in south-exposed wood blocks
In order to investigate whether the associations of WIF and N2-fixing bacteria might have contributed to the differential deadwood decomposition dynamics between the north- and the south-facing site, association networks were calculated. The number of fungal OTUs considered for network calculations was comparable at both slopes (south = 119 OTUs, north = 121 OTUs), as was the number of OTUs that were exclusively detected at each slope (Fig. 4A). A total of 122 N2-fixing bacterial OTUs annotated as Burkholderiales (49 OTUs) and Rhizobiales (73 OTUs) were used for network analyses (Fig. 4B). The richness of N2-fixing bacterial OTUs was higher in the south-exposed wood blocks (south = 118 OTUs, north = 69 OTUs). Except for four OTUs, all N2-fixing bacterial OTUs detected at the north-facing site were also found at the south-facing one (Fig. 4B).
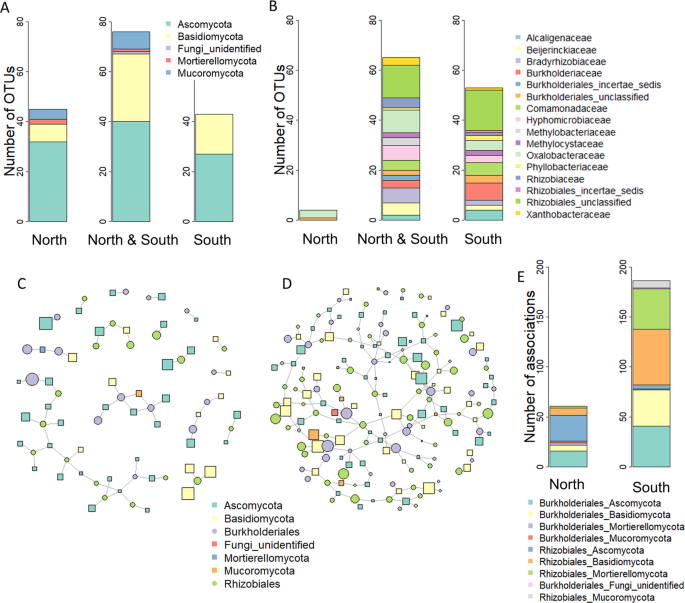
Associations between fungal and N2-fixing operational taxonomic units (OTUs) in the P. abies experimental wood blocks at the north- and the south-facing site. (A,B) Distribution of fungal (A) and N2-fixing bacterial (B) OTUs across the samples collected at north- and south exposure, respectively. The barcharts indicate the taxonomic distribution of the OTUs at the level of phylum (A) and family (B), respectively. Association networks calculated for the north- (C) and the south-facing (D) slopes. Every symbol represents a fungal or N2-fixing bacterial OTU. Lines indicate the association of two OTUs. Unconnected OTUs were not included into the model. Symbol sizes correspond to the abundances of the OTUs. The binary adjacency matrix of the calculated networks was visualized in R using package igraph62 (http://igraph.org). (E) The barchart illustrates the taxonomic annotations of the association pairs in the north- and south-exposed wood blocks.
As we hypothesized, the network of the south-exposed wood samples comprised a higher number of associations between fungal and N2-fixing bacterial OTUs (south = 168, north = 61, Table S3). Moreover, a higher percentage of fungal and N2-fixing bacterial OTUs were integrated into the south- (75% of all OTUs present) than into the north network (43%). A relatively small number of samples (12 for each slope) were used for network analysis, which might have resulted in a number of relevant associations undetected and some false positive associations between OTUs. This latter aspect is an inherent problem in network analyses38. However, these limitations were overcome by applying stringent filtering prior to calculation (Table S4) and using a sparse covariance matrix and strict thresholds in the algorithm. Furthermore, a high number of spurious associations were unlikely to occur considering the coherence of the network across a wide range of strict settings (Table S4). Indeed, we interpreted associations if they were only reoccurring in several differently calculated networks. Moreover, both north and south networks were composed of OTUs with high abundances rather than small OTUs (Table S3).
In addition to size and density, the respective fungal-bacterial associations may also determine the effect on deadwood decomposition. All of the OTU associations were slope-specific (Fig. 4C–E). At both north- and south-facing sites, a comparable number of fungal OTUs were associated to Burkholderiales and Rhizobiales OTUs (Fig. 4E, Table S3). In the south network 60% of fungal OTUs and 80% of OTUs characteristic for the later stages of decomposition (52 and 104 weeks) had more than one association with a N2-fixing bacterial OTU (Table S3). Of those OTUs, 70% of all OTUs and 77% of all indicator OTUs of the later stages showed at least one positive and one negative association in the south network (Table S3). This repetitive pattern emphasizes the ecological relevance of the interactions between fungi and N2-fixing bacteria in deadwood and pinpoints that fungi might influence the N2-fixing bacterial composition in their habitat.
Basidiomycota, mainly yeasts, that were dominant in terms of abundance at the earlier decomposition stages (Fig. 3A, Table S2) accounted for 41% and 21% of all the associations in the south and north networks, respectively (Fig. 4C–E, Table S3). Wood-decay fungi have been associated with members from the family Burkholderiaceae20, and co-occurrence between Burkholderiales and fungi was also reported for soil environments on a global scale39. The increased association to N2-fixing bacteria might have provided the basidiomycete fungi with a better access to nitrogen, which is a limited resource in deadwood, especially in the early stages of decomposition40, ultimately leading to a faster decay. The Basidiomycota with the highest abundance and strongest associations to Burkholderiales in the south network were the yeasts Guehomyces pullulans and Leucosporidiella creatinivora (= Rhodotorula creatinivora) (Table S3). These two species are saprobial generalists degrading plant litter and wood xylan41. L. creatinivora can degrade phenol and phenol-related compounds in cold habitats42, indicating that they could also be capable of degrading lignin. In addition, L. creatinivora also had positive associations with Rhizobiales (Table S3). These associations were not detected in the north network (Table S3). It is very likely that these associations provide additional competitive advantages for early wood colonization due to the potential N-enrichment of the substrate.
In the later stages of decomposition (52 and 104 weeks), there was an increase in the abundance of Ascomycota at both slopes (Fig. 3A). They were associated with Burkholderiales as well as Rhizobiales (Fig. 4E). In the south network, a strong positive association was found between the mycorrhizal Meliniomyces OTUs and both Burkholderiales and Rhizobiales (Table S3). These associations were absent in the north network (Table S3). Meliniomyces was the taxonomic group with strongest within-group interconnections (Table S3), which might further support the relevance of mycorrhizal fungi in the WIF community (Fig. 3C). Some associations between fungi and Burkholderiaceae strains are based on the co-migration of Burkholderiaceae strains with fungal hyphae43,44. This scenario, in which Burkholderiaceae provide protection from antifungal agents while feeding on fungal glycerol, thus supporting fungal growth, might be beneficial for deadwood decomposition.
In contrast thereto, in the north network, characteristic OTUs for the later sampling points with strong associations were Chlarala – Rhizobiales, Pseudogymnoascus – Rhizobiales and Mucor – Burkholderiales (Table S3). Chalara are anamorphic ascomycetes able to degrade cellulose45. However, this group also includes important phytopathogenic fungi, such as C. fraxinea causing ash dieback. Mucor was often associated with Burkholderiales bacteria (Table S3), and this so-called fungus-associated bacteriome is known to be important for the host’s lifestyle and interactions. Shifts in the composition of the fungal-associated bacterial community can slow fungal growth and change fungal secondary-metabolite production46.
Generally speaking, the higher number of associations between fungal and N2-fixing OTUs likely favoured the ecosystem performance, i.e. deadwood decomposition rate at the south-facing slope when compared to the north-facing one. This is in agreement with an emerging study from Wagg et al.47, which points towards the importance of microbial inter-kingdom associations as a driver of ecosystem functioning. They demonstrated that microbial networks with a greater number of associations between bacterial and fungal taxa contributed more to support multiple ecosystem functions simultaneously related to soil nutrient cycling than simpler or low-diversity networks.
Source: Ecology - nature.com
