The boratic microbialite, Shivlinga
Shivlinga is a 35-cm-tall microbialite that is several decades old according to residents of the closest village situated at the eastern end of the Puga valley (Fig. 1A,G). The abundance of microbial biomass on its surface and chemical composition of its mineral body (see below) suggest that Shivlinga has formed via precipitation of boratic minerals on lithifying microbial mats. Laminated, clotted or dendritic fabrics commonly found on stromatolites, thrombolites or dendrolites43 were not encountered on Shivlinga’s mineral deposits. It is for this reason that Shivlinga was considered to be a microbialite, i.e. a microbe-mediated sedimentary rock-like mineral deposit with or without defined internal fabrics43. The near-circular, flat top of Shivlinga’s mineral body has a mean diameter of ~22 cm (Fig. 1C,I). Throughout the year, there is a gentle discharge of water (68–73 °C, pH 7.0–7.5) from a 10-cm diameter vent at the summit of Shivlinga; this is accompanied by a weak emission of hydrothermal gases (a comprehensive characterization of the gaseous discharges of the geothermal vents of Puga valley has shown that they all emit steam, carbon dioxide and hydrogen sulfide41). Shivlinga’s discharge runs down the side of the microbialite (Fig. 1C,I) onto the bedrock surrounding its base (Fig. 1D,J). Beyond the slopes of the basement rock, the spring-water flows along several shallow channels (the longest one running 3.3 m from Shivlinga’s base), and eventually percolates into the regolith of the surrounding apron (Fig. 1B,H). Temperature, pH and flow-rates of the vent-water and outflows were found to remain consistent during the 2008–2013 annual site visits (Table 1 and Supplementary Table 1); this stability was attributable to the physicochemical consistency of the underlying geothermal reservoir44. The sizes and structures of the microbial mats growing along Shivlinga’s spring-water transit also remained largely unchanged during the period 2008–2013, indicating the microbiome’s resilience to seasonal and annual variations in weather. This observation was consistent with reports for microbial mats at other geothermal sites22,25; so Shivlinga’s ecosystem was considered suitable for studying microbial community dynamics along its hydrothermal gradients, and a comprehensive geomicrobiological exploration of the site was undertaken in July 2013.
Chemical characteristics of the vent-water
The chemical composition of Shivlinga’s vent-water was determined by collecting surficial discharges from the center of the vent’s orifice. The vent-water, at the time of the current sampling (on 23 July 2013), had a neutral pH (7.0) and a low concentration of total dissolved solids (2000 mg L−1) compared with other neutral-pH hot springs located in distinct geographical areas of the world41,45,46. Shivlinga’s vent-water was found to have high concentrations of boron (175 mg L−1), sodium (550 mg L−1), bicarbonate (620 mg L−1) and chloride (360 mg L−1), compared to the other solutes detected in the vent-water. There was also some silicon (60 mg L−1), potassium (15 mg L−1), calcium (10 mg L−1), lithium (6 mg L−1) and magnesium (3 mg L−1) detected. Of the sulfur species present, thiosulfate (3 mM) and sulfate (1 mM) were the most abundant, but sulfite (225 μM) and sulfide (250 μM) were also present at significant concentrations. Collectively, the vent-water chemistry was consistent with that reported for other hot springs within the Puga region40,41.
Distinct mineralogies of Shivlinga’s vent and body, bedrock slope, and apron
For each of the water-flows running in Shivlinga’s vent-to-apron trajectory, and starting from the surface of the vent-water, multi-colored microbial mats grow all along their transits, until the end of the flow near the edge of the apron. Depending on their thickness, the mats are either slightly submerged or stay just above the water level. Segments of mats that lie at the interface of the spring-water and regolith are typically dry at their surface but moist within. Every mat at the site is intermeshed with fresh mineral accretions that precipitate from the cooling spring-water and also condense from the gases that arise from the vent; but mineralization processes are at far more advanced stages in the less-hydrated segments of mats. For microbial biomass growing just beneath the water level, or protruding from the water surface by some mm to cm, mineral dusts visible to the naked eye are present on the mat surfaces. For those mats or parts of mats that are situated at the margins of outflows, mm- to cm-sized, soft, white spherules and shrub-like bodies of accreted minerals cover the surface (Fig. 2A). In the latter cases, mineralization is very conspicuous: monitoring this process over a period of 21 days using close-up photography revealed that the microbial mats grow out from the top of the encrustations, and spread again over the surface of the spherules (Supplementary Fig. 2). It was also evident that the fresh biomass at the surface of such mats are covered by fresh mineral deposition; and so the process continues. Mineralization acts to solidify all the mat structures of the microbiome from the bottom upwards. The microbialite thereby grows in height as well as in girth, besides hardening from within; the height of Shivlinga, according to our field observations, increased by ~4 cm (and the vent orifice narrowed slightly) between 2008 and 2013. Spherules and shrub-like structures form across the apron’s dry surface, presumably due to condensation of Shivlinga’s fumarolic gases on the regolith (Fig. 1B,H). However, the salt accretions which form beyond a few cm from the banks of the water-flows are not associated with microbial mats, and are dry and brittle.
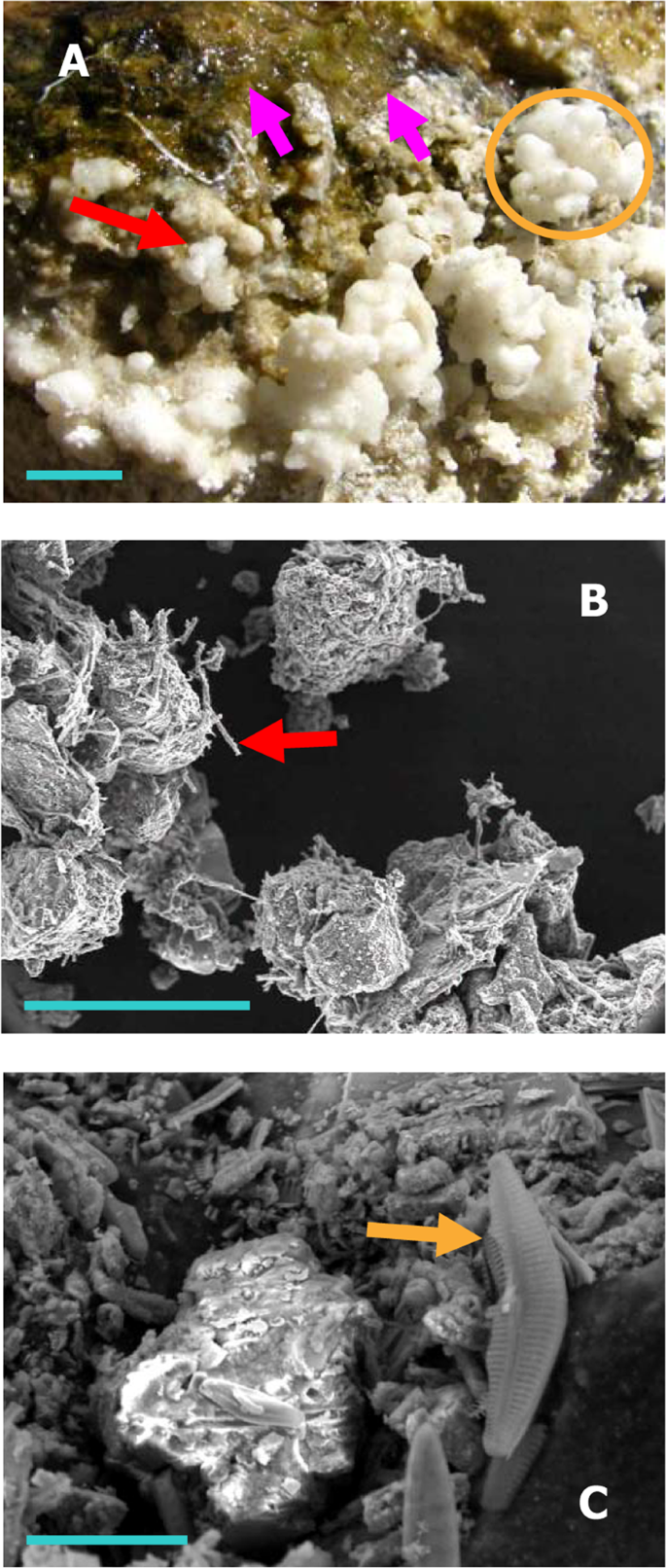
Macro-/micro-scale microbe-mineral structures at the Shivlinga site: (A) green microbial mat (pink arrows), and white spherules (red arrow) that collectively form shrub-like mineral bodies (orange circle), on the upper surfaces of the vent’s rim (blue scale bar = 10 mm); (B) sinter particles from 5-cm-deep inside the wall of the microbialite, associated with bacterial filaments (red arrow) (image was taken using SEM, blue scale bar = 150 µm); and (C) a diatom cell (golden arrow) associated with the particles shown in (B) (using SEM, blue scale bar = 20 µm).
We analyzed fine mineral particles accreted on the mat samples taken from the middle of the water-flow, and sinter-spherules precipitating on mat-portions growing near the margins of the water-flow. Sinter-spherules and shrub-like bodies that were soft, so had presumably formed within a few weeks before sampling, were collected from one top-edge of Shivlinga’s body (Sinter-Sample 1), one side-surface of the microbialite (Sinter-Sample 2), one point on the bedrock slope around Shivlinga’s base (Sinter-Sample 3), and one point at mat:regolith interface on the margin of the spring-water flow across the apron (Sinter-Sample 4) (Fig. 1B–D,H–J). Besides these four samples, older and harder sinter material was collected from Shivlinga’s interior by boring the microbialite’s side-surface, using a 3 mm twist drill bit, to a depth of 5 cm at a point located 15 cm below the summit (Sinter-Sample 5) (Fig. 1C,I).
All the sinter samples were found to be rich in boron, sodium and calcium (Supplementary Table 2); kernite (Na2B4O7·4H2O) was the major mineral in Shivlinga’s interior, while borax (Na2B4O7·10H2O) and then tincalconite (Na2B4O7·5H2O) were predominant in the soft, recently-formed sinters (data not shown). Calcite (CaCO3), gypsum (CaSO4·2H2O), elemental sulfur, and silica (SiO2) were also identified within the boron-mineral matrices of all the five sinter samples. Minute quantities of aluminum, gold, iron, manganese, zinc, molybdenum, lead, silver, nickel and cobalt were also detected, regardless of the sinter samples (Supplementary Table 2). These mineralogical data are consistent with Shivlinga’s vent-water chemistry, and together they suggest that the microbialite has formed from the gradual precipitation and accretion of boratic-, carbonatic-, and sulfatic-minerals, a process that, in part at least, is facilitated by its microbial communities (see Fig. 2A).
The process of precipitation of boron minerals is common to Shivlinga’s vent and microbialite body, bedrock slope and apron; yet there is considerable variation in the proportions of oxides or carbonates of metals, alkali metals and alkaline earth metals which combine with the boron minerals in the three zones of this microbiome (Supplementary Table 2). The soft (fresh) sinters of the vent and microbialite body, and those of the sloping bedrock, contain greater proportions of aluminum and gold, while sinters of the apron contain greater proportions of iron and manganese. On a w/w basis, the percentages of zinc, molybdenum, lead, silver, nickel and cobalt etc are higher in the freshly-precipitated sinters located within the apron than in the accretions on the vent and microbialite body, or the bedrock slopes (Supplementary Table 2).
Microbial characterizations along Shivlinga’s spring-water transit/dissipation: a wet, and a progressively-drying, thermal gradient
Microbiological investigations were carried out along two distinct axes of Shivlinga’s spring-water transit/dissipation. One of these represented a progressively-drying thermal gradient having no running water on it (along this axis, the spring-water dissipated into the mineral sinters of the microbialite body); the other featured a wet thermal gradient along the outflow channel of the spring. Both of these gradients start from the vent and, besides the vent-water community (VW), share a common microbial mat community (VWM) that floats on the vent-water and anchors to the sintered rim of the vent (Fig. 1C,I). The drying thermal gradient traverses the moist sinters concentrically from the vent opening, ~15 cm towards the edge of Shivlinga’s flat-topped summit; two more, morphologically distinct but physically contiguous mat communities (DG3 and DG4) occur along this trajectory (Fig. 1C,I). On the other hand, the ~4-m-long wet thermal gradient, at the time of sampling, laid along the longest and widest of the six prominent channels of Shivlinga’s spring-water transit; it ran from the vent, down one side of Shivlinga’s mineral body and base (Fig. 1C,D,I,J), and then along the outflow channel in the apron (Fig. 1B,H). Along the wet thermal gradient lies a continuum of multi-hued microbial mat communities, from within which five representative mats (WG3, WG4, WG5, WG6 and WG7) were chosen for investigation. In total, nine physically/morphologically distinctive microbial communities, including one in the vent-water (Table 1), were sampled across the two thermal gradients and subjected to analysis using microscopy and metagenomics. These microbiological data were collated with the mineralogical analyses of sinter materials (sediment-accretions) sampled and investigated from the five distinct depositional facies (physical and chemical conditions under which deposition takes place) existing in the Shivlinga territory. This revealed that the Shivlinga microbiome encompasses three distinct geomicrobiological zones (Fig. 1A,G) characterized by distinctive temperature- and pH-conditions, and topographical, mineralogical and microbiological features. These are (i) the vent and the microbialite body, (ii) the microbialite’s base (including the sloping bedrock) and (iii) the apron. Whilst the first geomicrobiological zone encompassed the VW, VWM, DG3, DG4 and WG3 communities (Fig. 1C,I), the second and the third included the communities WG4 and WG5 (Fig. 1D,J), and WG6 and WG7 (Fig. 1B,E,F,H,K,L), respectively. Further details of the geomicrobial features present along Shivlinga’s spring-water transit/dissipation are given in the Methods sub-section titled “Site Description”.
Distinct mat morphologies of the vent surface, microbialite body, sloping bedrock-base, and apron
Microscopic examinations revealed four distinct patterns of organization of microbial cells in Shivlinga’s mat communities. The VWM and WG3 streamers are composed of rod-shaped bacterial cells that lie end-to-end, interspersed with smaller coccoidal cells, within dense networks of aseptate filaments of cyanobacteria that are long, straight, sheathed and semi-rigid (Fig. 3A,B). All the green, red and purple mats growing around the vent, on the microbialite body, and the bedrock slope of Shivlinga (i.e. DG3, DG4, WG4 and WG5) are made up of spherical bodies ranging from 10–100 µm in diameter (Fig. 3C). Diatoms that appear to be members of Cymbellaceae form the margins of these spheres, and remain in immediate contact with adjacent spheres or other mineral structures (Fig. 3C,D). Cells, which have the dimensions and morphologies of prokaryotic cells, occur along the boundary of the spheres. Filamentous and coccoidal microorganisms, resembling Chloroflexi and Chroococcales respectively, occupy the internal core of the spheres; abundant hyphal structures that are aseptate and white (some have stalked buds coming out of them), and resemble Phycomycetian fungi are also seen here (Fig. 3E–H). Notably, boron mineral-encrusted forms of these spherical bodies were identified in SEM-EDS of Sinter-Sample 5 (Fig. 2B,C). These microscopic data, together with the detection of small but definite proportions of fungus-affiliated reads in the subsequent metagenome analysis of the mat communities, indicated that specific amplification and sequencing of fungal DNA, in future investigations, may reveal even greater fungal diversity than shotgun metagenomics indicates.
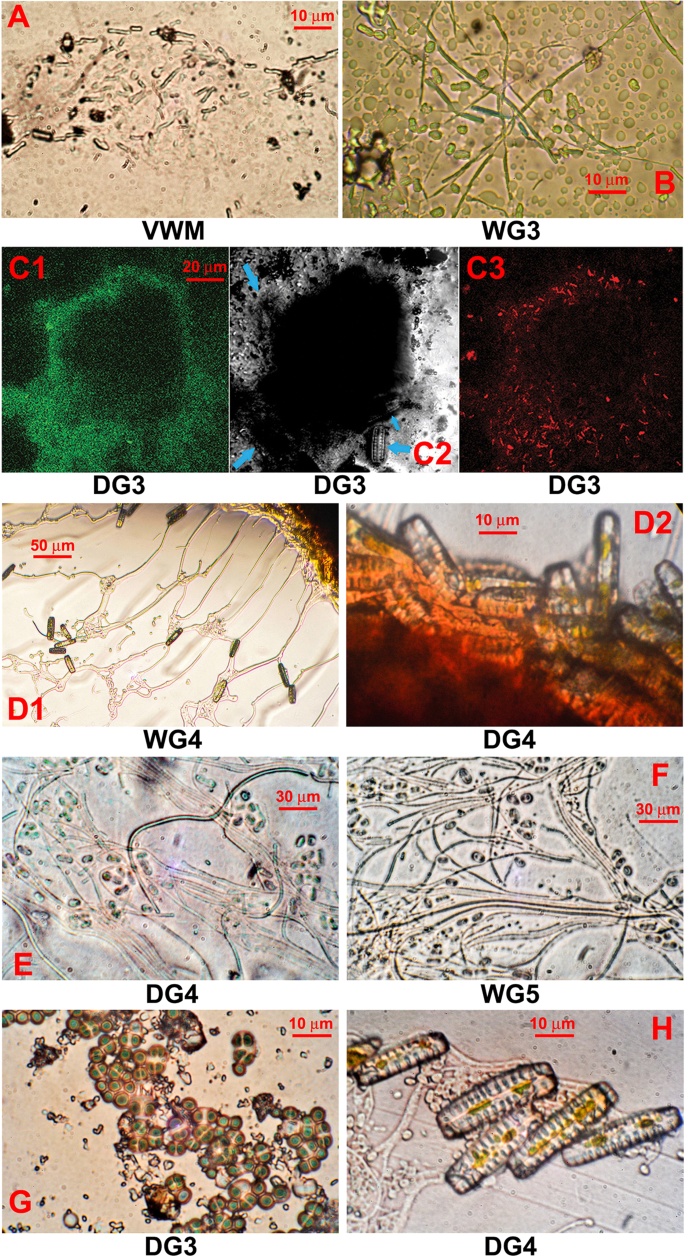
Microbial structures from the Shivlinga site: (A and B) microbial cell filaments from streamers of VWM and WG3, respectively; (C) microbial biomass of DG3; (D) diatoms from the surface of the microbial mass which makes up WG4 and DG4 (D1 and D2 respectively); (E and F) cells from the core of the microbial mass which makes up DG4 and WG5, respectively; (G) Chroococcales-like cyanobacteria in the core of the microbial mass which makes up DG3; (H) fungal hyphae and diatoms near the edge of the microbial mass which makes up DG4. All images were produced using phase contrast microscopy except for (C) that used laser scanning confocal microscopy. For (C1), excitation was carried out at 488 nm and detection at 630–650 nm; (C2) is a differential interference contrast image with blue arrows indicating diatom cells; and for (C3), excitation was carried out at 543 nm and detection at 650 nm.
The two morphologically distinct mat structures of the wet thermal gradient that are located within apron environment, i.e. WG6 and WG7, suggest processes by which mat formation had occurred. The former type appears to have come about via nucleation of borax, extracellular organic matter, and traces of calcite and other minerals, on irregular aggregations of diatoms, cyanobacteria, red-pigmented Chloroflexi, and other cells having dimensions and morphologies akin to those of prokaryotes. The WG7 streamers, in contrast, exhibited a laminar organization. The top-most layer of these stratified mats contained dense sheaths of filamentous bacteria resembling Chloroflexi, Leptothrix and Sphaerotilus. Embedded in complex mineral assemblages, these filamentous organisms are interspersed with large round-shaped cells that appear to be flagellates, according to their morphologies and 5–20 µm diameters (row A of Fig. 4). At 1–2 mm beneath the top-layer (i.e. in sub-surface layer 1), abundance of the filamentous bacteria decreases and the density of the eukaryotic cells increases; at these depths, there are also occasional cells of pennate diatom species (row B of Fig. 4). Diatoms are, however, much more abundant at subsequent depths up to the boratic substratum. At 3–4 mm from the mat surface (i.e. in sub-surface layer 2), the diatoms co-exist with morphologically-diverse bacteria, many of which resemble purple sulfur and non-sulfur bacteria (row C of Fig. 4). At the deepest layer of WG7, 5–8 mm below the surface (i.e. in sub-surface layer 3), diatom cells predominate and are attached to borax crystals (row D of Fig. 4).
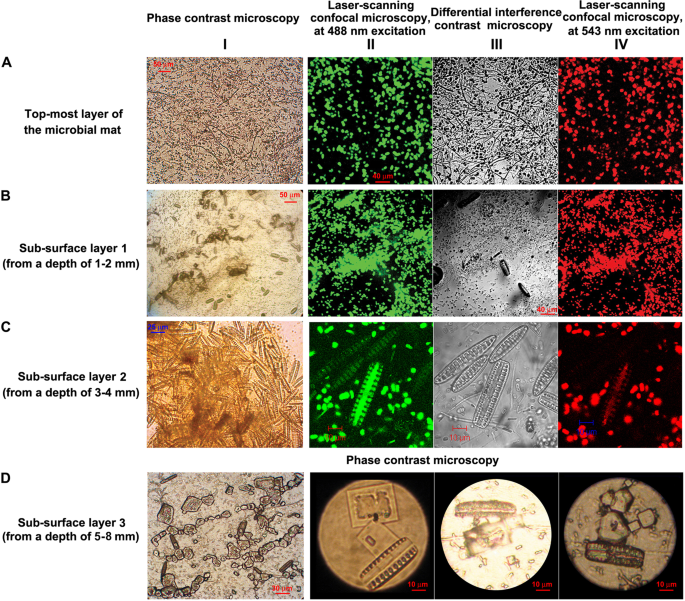
Microbial cells present in the different layers of the WG7 sample: (row A) top-most layer of the microbial mat; (row B) sub-surface layer 1 (from a depth of 1–2 mm); (row C) sub-surface layer 2 (from a depth of 3–4 mm); and (row D) diatom cells and borax crystals in sub-surface layer 3 (from a depth of 5–8 mm); (Column I) images were taken using phase contrast microscopy; (column II, except for row D) images were taken using laser scanning confocal microscopy, at 488 nm excitation and 630–650 nm detection; (column III, except for row D) used differential interference contrast to modify the image from column II; and (column IV, except for row D) used laser scanning confocal microscopy (543 nm excitation and long pass 650 nm detection) to further modify the image from column II. Photographs in row D were taken using phase contrast microscopy.
Bacteria-dominated vent-water community
PCR amplification (using Bacteria-/Archaea-specific oligonucleotide primers), followed by high throughput sequencing, of the V3 regions of all 16S rRNA genes present in the total environmental DNA extracted from the VW sample revealed the alpha diversity of the community. This is referred to hereafter as the metataxonomic composition47 so as to distinguish the diversity reported for the VW community from the diversities reported subsequently for the mat communities on the basis of shotgun metagenome sequencing and analysis. Notably, total environmental DNA yield from the VW sample was insufficient for direct shotgun sequencing and metagenome analysis.
Bacteria-specific V3 primers generated PCR products of desired size (~200 bp), so the amplification product was sequenced at high data throughput. Archaea-specific V3 primers did not yield any PCR product, suggesting that very low numbers of archaeal cells are present in Shivlinga’s vent-water. Reads of the Bacteria-specific V3 sequence dataset were clustered into operational taxonomic units (OTUs) or putative species-level entities unified at the level of 97% 16S rRNA gene sequence similarity (Supplementary Table 3 shows the summary results of OTU clustering). Rarefaction analysis of the dataset confirmed that the read-sampling level (data throughput) achieved was sufficient to reveal most of the diversity present in the sample (Supplementary Fig. 3).
Total 64 bacterial OTUs – distributed over the phyla Proteobacteria (45), Actinobacteria (4), Firmicutes (4), Deferribacteres (2), Deinococcus–Thermus (2), Ignavibacteriae (2), Armatimonadetes (1), Aquificae (1), Synergistetes (1) – were identified in the VW sample (number of OTUs affiliated to each phylum is given in parenthesis); 2 of these belonged to unclassified Bacteria. Of the 64 OTUs, 19 could be classified at the genus level. The genera identified (number of affiliated OTUs given in parenthesis) were Achromobacter (2), Alcaligenes (1), Aminicenantes Incertae Sedis (1), Armatimonadetes gp5 (1), Calditerrivibrio (2), Ignavibacterium (2), Paenibacillus (1), Propionibacterium (1), Sulfurihydrogenibium (1), Thermomonas (4), Thermus (2) and Thiofaba (1). Notably, out of the 12 genera detected metataxonomically in the 70 °C VW, member strains of at least four have never been found to grow at >45 °C in vitro (these are Achromobacter, Alcaligenes, Paenibacillus and Propionibacterium); and six are present in each of the mat communities studied (these are Achromobacter, Calditerrivibrio, Paenibacillus, Propionibacterium, Sulfurihydrogenibium and Thermus).
Microbiology of Shivlinga’s thermal gradients
Variations in the community composition of Shivlinga’s microbial mats were assessed via metagenome sequencing and analysis along each thermal gradient (Supplementary Tables 4 and 5 show the summary statistics plus domain-level classifications of the metagenomic data obtained from the mats of the drying and wet thermal gradients respectively). Although total number of metagenomic reads generated for the different samples varied, plateauing of rarefaction curves determined for the individual samples by plotting number of reads analyzed versus genera identified in searches against the non-redundant (nr) protein sequence database of National Center for Biotechnology Information (NCBI, USA) or the 16S rRNA gene sequence database of the Ribosomal Database Project (RDP) showed that the data throughput achieved was sufficient to reveal most of the diversity present in the sample. Furthermore, to avoid the risk of potential anomalies that may arise in the subsequent analyses due to differential read-count of the different samples, prevalence of taxa was quantified as the percentage of total metagenomic reads ascribed to them, i.e. relative abundance.
In searches against the nr protein database, unassigned and unclassified sequences made up 7–26% of the metagenomic readsets; Bacteria accounted for at least 72% of all these and Archaea constituted <1%, regardless of the sample. The highest relative abundance of Archaea was encountered in the distal (cooler) ends of the gradients, i.e. in DG4 (41 °C), WG4 (46 °C) and WG7 (33 °C) communities. The genera Archaeoglobus, Methanocaldococcus, Methanococcus, Methanosarcina, Methanospirillum, Methanothermobacter, Pyrococcus and/or Thermococcus comprised the major archaeal component of every mat community. Eukarya accounted for <0.6% of the metagenomic readsets for VWM, WG3 and WG4; and ~1% for DG3, DG4; WG5, WG6, and WG7 (for at least one replicate). In other words, relative abundance of eukaryotes almost doubled along the drying and wet thermal gradients at ≤52 °C and ≤38 °C (respectively), compared to those observed at higher temperatures. Diverse types of virus are present Shivlinga’s mat communities and, remarkably their relative abundance is highest at 66 °C (in VWM). Virus-affiliated reads made up 0.3% and 0.2% of the metagenomes obtained from the two VWM sample-replicates, but only 0.01–0.04% of those obtained from the other mat communities. Predominant members of the viral component of VWM metagenomes were temperate lactococcal phages, including r1t and Listeria phi-A118; notably, thermophilic phages that are commonly found in other hydrothermal ecosystems48 were absent. Whereas the Bacteria are the dominant organisms in Shivlinga’s microbiome, the ecophysiological importance of eukaryotic microbes could be greater than suggested by their low number of metagenomic reads. Some indications of this are detailed below.
Microscopic analyses revealed dense but localized populations of diatoms in many of the mat communities, including those located at 52 °C (Fig. 3C,D). Diatoms (Bacillariophyta) were also detected metagenomically in all of the microbial mats sampled, albeit in low numbers. Their relative abundance ranged from a minimum of 0.001%, for VWM, to a maximum of 0.05%, for DG3, along the drying thermal gradient and 0.6%, for WG5, along the wet thermal gradient (relative abundances values for diatoms were 0.2% for DG4, WG6 and WG7). The genera Odontella, Phaeodactylum and Thalassiosira consistently predominate diatom populations in all the eight mat communities, regardless of the hydrothermal gradient.
Microscopy revealed localized but dense populations of fungi in mat communities present at the 33–52 °C sites. According to metagenomic analyses, fungi were also present in all of the other mat communities, albeit at low levels. Along the drying thermal gradient, their relative abundance ranged from a minimum of 0.02%, for VWM, to a maximum of 0.2%, for DG4, and along the wet thermal gradient from 0.02% (VWM) to 0.7%, for WG5. Furthermore, 63 fungal genera were found to be present in VWM (66 °C), while greater numbers of genera were detected in the communities growing at the distal ends of each gradient; e.g. 82 genera in DG4, 106 in WG5, and 142 in WG7. Aspergillus, Gibberella, Neurospora, Saccharomyces, Schizosaccharomyces and Ustilago constituted the major portions of the fungal populations across both the gradients.
Sequences matching Chlorophyta (green algae) were present in the metagenomes of all the mat communities, and their relative abundance ranged from 0.005% (for VWM) to a maximum of 0.1% (for DG4, WG5, WG6 and WG7). The genera Chlamydomonas, Chlorella, Micromonas, Ostreococcus and Volvox are the major green algal component of all the mat communties, whereas Acetabularia, Bryopsis, Dunaliella, Nephroselmis, Pyramimonas and Scenedesmus, sparse at high temperatures, increase towards the distal ends of the gradients. Remarkably, the eukaryotic components of each mat metagenome were not dominated by a microorganism, but by the bryophyte Physcomitrella (spreading earthmoss) which has a simple life-cycle, and is considered to be the most-ancient of all land plants49. It is an early colonizer of exposed mud/ sediment/ soil around the edges of water bodies, and is widely distributed in temperate regions of the world. At the Shivlinga site, Physcomitrella is most abundant in VWM (66 °C), where it constitutes 0.1% of the metagenomes. Notably, in the VWM metagenomes, known thermophilic genera such as Archaeoglobus, Calditerrivibrio, Desulfotomaculum, Hydrogenivirga, Pyrococcus, Thermoanaerobacter and Thermocrinis, each has <0.1% of reads ascribed to them. In the other mat communities, Physcomitrella constituted 0.02–0.04% of the metagenomes. In VWM, the most prevalent eukaryote after Physcomitrella is another photosynthetic organism, Cyanidioschyzon which is a unicellular, ~2 μm-long red alga that often occurs in sulfur-containing, highly acidic hot springs at around 45 °C50. For all other mat communities, the second-most prevalent eukaryote is the amoeba Dictyostelium which is found in soils, predates bacteria, and is commonly known as slime mold51. The other protists detected in Shivlinga’s mat communities included the flagellate cryptomonad alga Guillardia, the glaucophyte Cyanophora, and the chloroplast-bearing amoeba Paulinella. Notably, all these genera were present in sample sites of ≤46 °C.
Along the drying, as well as the wet, thermal gradients (Fig. 5A; Supplementary Fig. 4 and Fig. 5B; Supplementary Fig. 5), mean relative abundances of the 26 major groups of Bacteria (i.e. 21 phyla and five proteobacterial classes) varied considerably. Deinococcus–Thermus and Aquificae dominate VWM, and are less prevalent in the communities located at the distal (cooler) ends of each gradient. Along the drying thermal gradient, this decline in the prevalence is very sharp for both phyla; relative abundance of Deinococcus–Thermus declined from 60% in VWM to ~2% in DG3 and DG4. In the drying thermal gradient, there was a concurrent increase of Chloroflexi; Firmicutes; Alpha, Beta, Gamma and Delta classes of Proteobacteria; Actinobacteria; Acidobacteria and Planctomycetes, which continues through DG3 and DG4. Notably, Cyanobacteria, Chlorobi, Bacteroidetes, Spirochaetes and Verrucomicrobia are more abundant in DG3 (relative to VWM), but declined in DG4 (relative to WG3). Contrary to the above trends, Epsilonproteobacteria and Thermotogae were low in DG3 (relative to their VWM levels) but increased again in DG4.
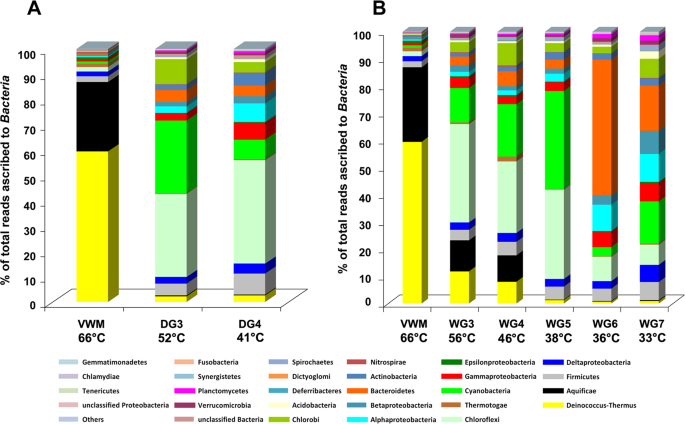
Mean percentage of reads ascribed to bacterial phyla, and classes within the Proteobacteria, in the duplicate metagenomes obtained from each mat community of (A) the drying thermal gradient and (B) the wet thermal gradient. The 21 phyla and five proteobacterial classes represented account for >0.1% reads in at least one of the 16 metagenomes analyzed. The category ‘others’ encompasses the phyla that accounted for <0.1% reads in every metagenome analyzed. Statistical significance of the fluctuations in the relative abundance of the taxa along the hydrothermal gradients can be seen in Supplementary Figs. 4 and 5 where their mean relative abundance within each mat community has been plotted alongside the two original relative abundance values obtained from the duplicate metagenomes (shown as vertical range bar in Supplementary Figs. 4 and 5).
Phylum-level population fluctuations are more irregular along the wet thermal gradient; the relative abundances of Alphaproteobacteria, Chlamydiae and Planctomycetes continue to increase along the water transit, in the vent-to-apron direction. The frequencies of Actinobacteria, Cyanobacteria and Spirochaetes also increase up to the 38 °C site. Similarly, Acidobacteria, Bacteroidetes, Chlorobi, Deferribacteres, Deltaproteobacteria, Dictyoglomi, Firmicutes, Fusobacteria, Gemmatimonadetes, Nitrospirae, Synergistetes and Tenericutes increase up to the 46 °C site. By contrast, the frequencies of Aquificae and Epsilonproteobacteria continue to decrease along the gradient, down to the 38 °C sample site, and Deinococcus–Thermus decreases down to the 36 °C site. Frequencies of Betaproteobacteria, Chloroflexi, Gammaproteobacteria and Verrucomicrobia increase till the 56 °C site. Remarkably, the relative abundance of Thermotogae does not vary with temperature along the wet thermal gradient. Consistent with these trends, only Aquificae, Deinococcus-Thermus and Epsilonproteobacteria exhibited significant positive correlation with both temperature and flow-rate of the spring-water (Supplementary Table 6). Whereas correlation of these three phyla with pH was significantly negative, only Aquificae showed significantly negative correlation with distance from the vent. Actinobacteria, Alphaproteobacteria, Chlamydiae, Firmicutes, Fusobacteria, Gammaproteobacteria, Gemmatimonadetes, Other phyla, Planctomycetes, Spirochaetes, Unclassified Bacteria and Unclassified Proteobacteria, all exhibited significant negative correlations with both temperature and flow rate. All these groups, remarkably, had significant positive correlations with pH and distance from the vent. Acidobacteria, Bacteroidetes, Betaproteobacteria, Chlorobi, Chloroflexi, Cyanobacteria, Deferribacteres, Deltaproteobacteria, Dictyoglomi, Nitrospirae, Synergistetes, Tenericutes, Thermotogae and Verrucomicrobia showed no significant correlation with temperature, although Acidobacteria, Betaproteobacteria, Deltaproteobacteria, Synergistetes and Verrucomicrobia had positive correlations with pH as well as distance from the vent – of these, Acidobacteria and Betaproteobacteria additionally exhibited significant negative correlations with flow rate. Tenericutes had significant positive and negative correlations with pH and flow rate respectively. Supplementary Tables 7–12 show the calculations for all the correlation coefficients and their Benjamini-Hochberg-corrected P values.
Thermodynamic constraints on microbial colonization
Thermodynamic constraints on the microbial colonization of Shivlinga sites were identified based on relative abundance of taxa along the hydrothermal gradients. These, in turn, were derived from the percentage of metagenomic reads ascribed to individual taxa within each community. Diversity within Shivlinga’s microbial communities increases towards the distal ends of the thermal gradients, and community composition changes from Aquificae/Deinococcus–Thermus– to Chlorobi/Chloroflexi/Cyanobacteria-dominated, and eventually to a Bacteroidetes/Proteobacteria/Firmicutes-dominated one (Fig. 5). Although the VWM community on Shivlinga’s vent-water is biomass-dense, its microbial diversity is lower than that of the mat communities located at the distal end of each thermal gradient. This is likely due to the habitability barrier imposed by the high vent-water-temperature that, in turn, is exacerbated by the low atmospheric pressure. At an altitude of 4438 m, atmospheric pressure is ~6.4 KPa, which is close to the 2.5–5 KPa threshold that is known to prevent in vitro growth of bacteria adapted to atmospheric pressure at sea level (i.e. ~101.3 kPa)52. Low pressure not only reduces the boiling point of water but can also contribute to the entropic destabilization of biomacromolecular systems (conversely, high pressures appear to mitigate against the chaotropicity of MgCl2 in deep-sea brine systems5). Conditions at the Shivlinga site, therefore, destabilize biomacromolecules, and elicit cellular stress responses which impose high energetic cost on microbial systems and/or can ultimately cause cell-system failure53,54. For VW and VWM (the communities at the highest temperatures) an environment-driven selection for thermotolerant/thermophilic taxa would be expected. Accordingly, relative abundance of 19 out of the 26 major bacterial phyla/proteobacterial classes present along the wet thermal gradient was lower in VWM (66 °C) than in WG3 (56 °C). Only Aquificae, Deferribacteres, Deinococcus–Thermus, Dictyoglomi, Epsilonproteobacteria, Nitrospirae and Thermotogae, which are made up mostly of thermophiles, were more prevalent in VWM than in WG3 (Fig. 5B). This is indicative of a substantial thermodynamic barrier to microbial colonization at ~60 °C. The 19 groups, which include Alphaproteobacteria, Betaproteobacteria and Gammaproteobacteria, Bacteroidetes, Chlorobi, Chloroflexi, Cyanobacteria, and Verrucomicrobia, and for which prevalence was lower at 66 °C than at 56 °C, are made up mostly of mesophilic or thermotolerant members incapable of laboratory growth at >60 °C. The presence of these taxa at in situ temperatures of >60 °C, suggests that (a) hitherto unidentified environmental factor(s) is (are) acting to mitigate the cellular stresses induced by high temperature. Furthermore, the relative abundances of most of the major phyla/classes (20 out of 26) were lower at 56 °C, in WG3, than at 46 °C, in WG4 (Fig. 5B; Supplementary Fig. 5). This suggests that temperatures between 56 °C and 46 °C impose an additional thermodynamic barrier to habitability for many of the microbial taxa. In microbiomes for which habitability of some taxa is constrained by other parameters, a comparable drop in diversity is observed for xerophiles and halophiles at 0.720 water activity55. Furthermore, at the distal end of the wet thermal gradient, the relative abundances of 16 out of the 26 major groups were lesser at 46 °C (WG4) than at 38 °C (WG5), implying that there is another colonization barrier at 40–42 °C.
Simpson Dominance (D), Shannon–Wiener Diversity (H), and Shannon–Wiener Evenness (EH) Indices were calculated based on the metagenomic data for each mat community (Supplementary Tables 13–20); these values were then compared along each gradient. The trends revealed provide further evidence for temperature barriers to habitability. For VWM (66 °C), the D value was greater than for any other mat community, whereas H and EH were low relative to those for WG3 and DG3 (56 and 52 °C respectively) (Fig. 6). This indicates that, for both the wet, and the drying, thermal gradients, there is a colonization-barrier for many taxa at ~60 °C. However, along the wet thermal gradient, from 56 to 46 °C (WG3 to WG4), and along the drying thermal gradient, from 52 °C to 41 °C (DG3 to DG4), increases in H and EH, were much lower than those recorded in the respective gradients, across the 60 °C barrier (Fig. 6). This indicated that colonization-barriers at lower temperatures (~50 °C for the wet thermal gradient and 46–50 °C for the drying thermal gradient) are weaker than that at ~60 °C. Trends of the above indices further revealed that below 46 °C, along the wet thermal gradient, diversity did not correlate with temperature, so other biotic and/or abiotic parameters could be acting as determinants of community composition in this territory.
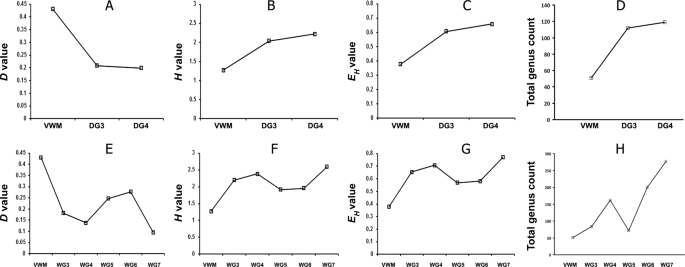
Simpson Dominance (A,E), Shannon Diversity (B,F) and Shannon Equitability (C,G) Indices, and total genus count (D,H), along the drying (A–D), and the wet (E–H), thermal gradients.
For each mat community, the most-abundant genera that collectively accounted for ≥50% of all classifiable reads were identified from the results of the metagenome-searches against nr protein database. The number and identities of such genera were then compared along the thermal gradients to interpret the temperature-constrained community dynamics. One genus, Thermus, accounted for a mean 50.6% of VWM metagenomes, and the genera Sulfurihydrogenibium, Meiothermus and Aquifex constituted 21.6%, 2.2% and 2.1%, respectively. For the DG3 and DG4 communities, 50% of their metagenomic reads were made up of 12 and 50 genera, respectively (Supplementary Table 21), and for WG3, WG4, WG5, WG6 and WG7, 50% of reads represented one, 11, 17, 35, 68 and 81 genera, respectively (Supplementary Table 22). In order to confirm the identifications of genera present in the individual mat communities, genus-level analysis of their duplicate metagenomes was carried out by searching against the 16 S rRNA gene sequence database of RDP. Using this approach (which is distinct from the amplified 16 S rRNA gene sequence-based metataxonomic approach, used only for the vent-water community), diverse bacterial, but no archaeal, genera were identified in all the eight mat communities (Supplementary Tables 23–30). The numbers of genera detected in this way varied along the two thermal gradients, and were consistent with the variations in microbial diversity indices (Fig. 6). For the drying thermal gradient, genus count was considerably less at 66 °C than at 52 °C or 41 °C (Fig. 6D). For the wet thermal gradient, genus count was far less at 66 °C than at 56 °C or 46 °C; unexpectedly, however, genus count was higher at 46 °C than at 38 °C, even though it was much lower at 38 °C than at 36 °C or 33 °C (Fig. 6H). The trends in microbial diversity that were revealed from the direct classifications of the metagenomic reads were consistent with the results of diversity analyses based on assembly of contigs and binning of population genomes from the mat metagenomes (Supplementary Note 2).
Key metabolic attributes of the Shivlinga mat communities
Analysis of the metagenomic data of individual mat communities for comparative richness of genes [or Clusters of Orthologous Groups (COGs) of Proteins] under various metabolic/ functional categories was followed by hierarchical clustering of the mat communities in terms of their enrichment of various COG categories. This revealed a dichotomy between DG3, DG4 and WG5 on one side, and VWM, WG3, WG4, WG6 and WG7 on the other (Fig. 7A). DG4 and WG5 clustered on the basis of their similarities with respect to high/low presence of COGs affiliated to the categories Cell motility; Replication, recombination and repair; Signal transduction mechanisms; and Translation, ribosomal structure and biogenesis. DG3 joined this cluster based on significantly high and low presence of COGs affiliated to Signal transduction mechanisms and Cell motility, respectively (Fig. 7B). In the other major cluster, the closeness of VWM and WG3 (Fig. 7A) is explained by their similarities in having high and low presence of COGs affiliated to the categories Replication, recombination and repair, and Signal transduction mechanisms, respectively (Fig. 7B). WG6 joined the VWM-WG3 cluster on the basis of significantly high presence of COGs affiliated to Replication, recombination and repair (Fig. 7B). WG4 and WG7, in turn, associated on the basis low presence of COGs for Replication, recombination and repair. VWM, WG3, WG4 and WG6 were further unified by the significantly low presence of COGs affiliated to Signal transduction mechanisms. Despite the significantly low presence of COGs affiliated to Signal transduction mechanisms in a number of mat communities, number species-level matches as well as relative abundance for genes encoding bacterial two component kinases such as histidine kinase, serine threonine protein kinase, diguanylate cyclase and PAS sensor protein were considerably high for all the communities. This implied that the geothermal adaptations of complex microbial mat communities involve efficient response regulations to a wide range of environmental signals.
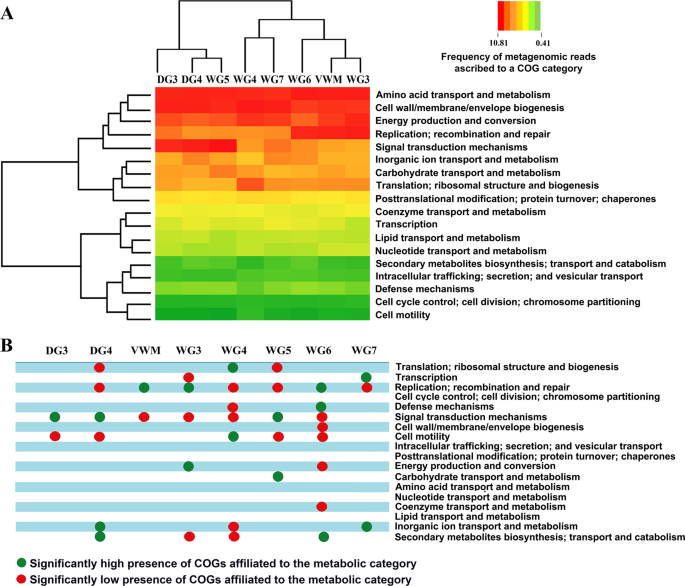
Functional analysis of the metagenomes isolated from the eight microbial mat communities of Shivlinga: (A) heat map comparing the richness of the metabolic/functional categories across the communities, determined in terms of the number of Clusters of Orthologous Groups (COGs) of Proteins that are ascribed to the categories in individual communities; a two-dimensional clustering is also shown, involving the eight mat communities on one hand and the 18 functional categories of COGs on the other; color gradient of the heat map varied from high (red) to low (green), through moderate (yellow), richness of the categories across the communities; (B) Statistically significant high (green circles) or low (red circles) richness of the functional categories across the communities, as determined by Chi Square test with p < 0.001.
The merged metagenomic readsets of individual mat communities were searched for genes putatively encoding reverse gyrase enzymes, which are typical of hyperthermophilic bacteria and archaea, but never found in mesophilic microorganisms56. As expected, metagenomic reads matching reverse gyrases from 30 different bacterial species were identified in VWM, but in no other community of the drying thermal gradient. Along the wet thermal gradient, such reads were found exclusively in communities living at ≥46 °C.
Metagenomic reads matching genes that encode key enzymes of the autotrophic Calvin–Benson–Bassham and Wood–Ljungdahl pathways, namely ribulose 1,5-bisphosphate carboxylase large chain (rbcL) and acetyl-coenzyme A synthetase (acsA), were identified in all the mat communities. Even as autotrophy is ubiquitous throughout the ecosystem, Wood–Ljungdahl pathway is apparently utilized by majority of the community members across Shivlinga’s thermal gradients. The numbers of species-level matches identified for acsA-related reads were an order of magnitude greater than those for rbcL; across the communities, species-level matches for acsA ranged between 112 (in WG5) and 477 (in WG7) while those for rbcL ranged from 18 (in DG4 and WG5) to 54 (in WG7). Furthermore, metagenomic reads corresponding to genes encoding the large subunit of ATP citrate lyase (AclA), a key enzyme of the reverse tricarboxylic acid (rTCA) or Arnon–Buchanan cycle – which is more energy-efficient, oxygen-sensitive, and phylogenetically ancient than the Calvin cycle or the Wood–Ljungdahl pathway57,58 – were detected only in the communities of the higher-temperature sites having typically low in situ oxygen, i.e. VWM and DG3 along the drying thermal gradient, and VWM, WG3 and WG4 along the wet thermal gradient.
Metagenomic reads matching genes for the gluconeogenic enzyme phosphoenolpyruvate carboxykinase (PEPCK) were detected in all the mat communities. Number of species-level matches for the PEPCK-related reads increased steadily from 32 in VWM to 102 in DG4, along the drying thermal gradient; and from 32 in VWM to 175 in WG7, along the wet thermal gradient. PEPCK governs the interconversion of carbon-metabolites at the phosphoenolpyruvate–pyruvate–oxaloacetate junction of major chemoorganoheterotrophic pathways, thereby controlling carbon flux among various catabolic, anabolic, and energy-supplying processes59. Variations in the number of species-level matches for PEPCK-related reads, therefore, were considered to be reflective of a steady rise in organoheterotrophic inputs in community productivity along both the hydrothermal gradients.
In view of the considerable abundance of dissolved sulfide, elemental sulfur, thiosulfate and sulfate in Shivlinga’s vent-water, occurrence and phylogenetic diversity of two key sulfur-chemolithotrophic genes, namely, the thiol esterase-encoding soxB of bacteria and the thiosulfate:quinone oxidoreductase-encoding tqoAB of archaea60, were investigated in the metagenomes of all the mat communities. While tqoAB was not present in any metagenome, soxB was detected in all of them. Number of species-level matches for soxB-related reads increased steadily from 4 in VWM to 16 in DG4, along the drying thermal gradient; and from 4 in VWM to 39 in WG7, along the wet thermal gradient. Sox-based sulfur-chemolithotrophs being predominantly aerobic60, these trends were consistent with the increase in oxygen tension along the vent-to-apron trajectories of the hydrothermal gradients.
Metagenomic reads matching genes encoding cytochrome c oxidase subunit 1 (cox1), the key enzyme of aerobic respiration61, are widespread in the Shivlinga ecosystem. Number of species-level matches for cox1-affiliated reads increased progressively from 32 in VWM to 193 in DG4, along the drying thermal gradient; and from 32 in VWM to 267 in WG7, along the wet thermal gradient. Metagenomic reads matching genes encoding dissimilatory sulfite reductase alpha and beta subunits (dsrAB), which is central to the energy-conserving reduction of sulfite to sulfide in anaerobic sulfite/sulfate-reducing prokaryotes62, were detected only in some of the mat communities. The number of species-level matches for dsrAB reads were 6, 0 and 3 in VWM, DG3 and DG4, respectively; and 6, 9, 17, 0, 2 and 16 in VWM, WG3, WG4, WG5, WG6 and WG7, respectively. No gene for N-oxide (NO/N2O) or nitrate (NO3) reduction was found in any mat community. Collectively, these data suggest that Shivlinga’s mat communities are adept in utilizing whatever low oxygen is there in their watery to semi-watery habitat. This hypothesis is further supported by the considerable occurrence and diversity (across the thermal gradients) of genes involved in the biogenesis of cbb3 cytochrome oxidase, which is known to be instrumental in respiration under very low oxygen tension63. For instance, the number of species-level matches for metagenomic reads ascribable to genes encoding the cbb3 cytochrome oxidase biogenesis protein CcoG64 increased progressively from 0 in VWM to 44 in DG4, along the drying thermal gradient; and from 0 in VWM to 87 in WG7, along the wet thermal gradient.
Source: Ecology - nature.com
