Review of the raw documentary video footage from the 2015 expedition generated three new fish species records for Rangitāhua: Labroides dimidiatus, Chaetodon mertensii and Ephinephalus rivulatus (Fig. 1). These species are considered tropical (or subtropical in the case of E. rivulatus21), consistent with previous studies that suggest a characteristic warm-water component in the fish fauna of Rangitāhua15,32,33,34,35. The observations of the cleaner wrasse, L. dimidiatus, and the yellow-back butterflyfish, C. mertensii from Rangitāhua represent the first records of these coral reef species for New Zealand waters. Both are cosmopolitan tropical Indo-Pacific species with their closest populations to Rangitāhua occurring at Teleki Tonga (South Minerva Reef) about 900 km to the northeast. Chaetodon mertensii also occurs at Norfolk Island about 1,300 km west of Rangitāhua36. Thus, our new records are extensions to the known geographic distributions for these fishes. The half-moon grouper, E. rivulatus is widespread throughout hard and soft bottom habitats of the Indo-West Pacific with its eastern distributional limit reported to be northeast North Island, New Zealand, Norfolk Island, New Caledonia and southern Japan22. This record of E. rivulatus from Rangitāhua extends its range eastward by about 700 km. All new records were of mature individuals, making it unlikely that these species represent new arrivals; instead, it is more likely that they were overlooked due to their rarity or residence in unsurveyed sites.
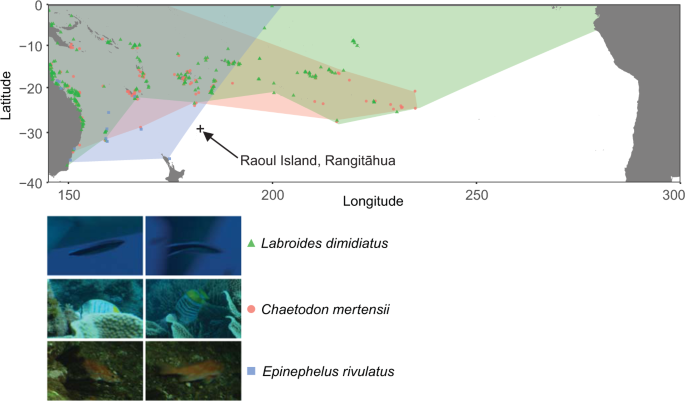
New species records for Rangitāhua. Species range maps36 and images (©NHNZ) of the new species records for Rangitāhua taken from the raw video footage. Raoul Island, Rangitāhua, is denoted by the cross and is shown to indicate the magnitude of the change in the known ranges for each species.
The documentary video sampled the fish assemblages for the shortest time period, and accordingly, captured the fewest number of species (see Supplementary Fig. S3). However, after accounting for variation in the duration of each survey, species richness captured in the documentary video dataset was lower than that observed by the timed transect and timed swim (t195 = 5.683, P < 0.001 and t195 = 5.230, P < 0.001 respectively; Fig. 2) but comparable to that observed by the timed stationary method (t195 = 0.019, P = 0.969, Fig. 2). Most of the reef footage in the documentary video we examined was stationary, thus it is likely that that the lower species richness of the timed stationary dataset and the documentary video dataset, was due to sampling a smaller area of reef and/or sampling a lower diversity of habitats than the swimming surveys of the timed swim and timed transect. Such stationary surveys are known to record lower fish species diversity, particularly for sedentary fish species37. Furthermore, even in the case of moving reef footage, estimates of species richness from video footage are typically lower than in visual census, due to field of view and difficulty in identifying fishes based on video footage alone38.
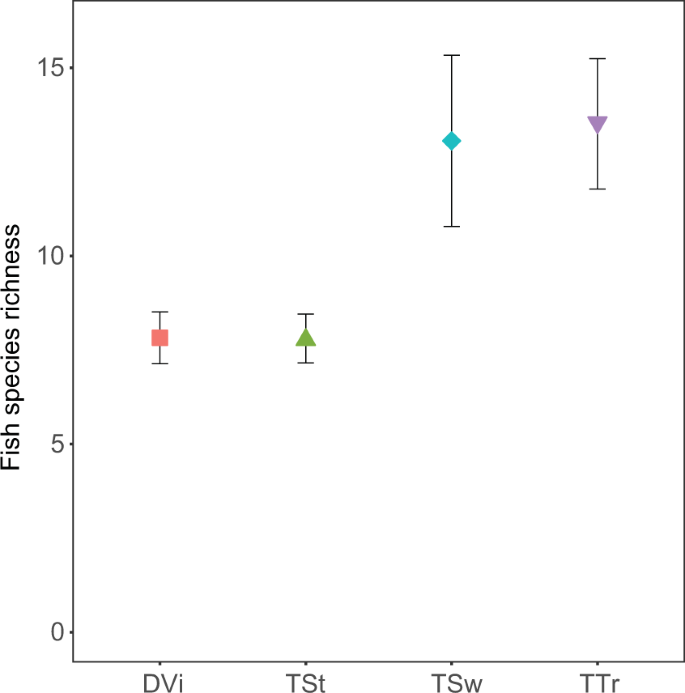
Surveyed fish species richness of Raoul Island, Rangitāhua for the average survey duration of 04:28 (mm:ss). Mean (±SE) species richness of fishes captured by the four different survey methods considered in our study (DVi = Documentary Video, TSt = Timed Stationary, TSw = Timed Swim and TTr = Timed Transect).
There were significant differences in the species composition observed among the surveys (Pseudo-F3,195 = 7.0475, Pperm < 0.001) and according to the time period surveyed (Pseudo-F1,195 = 1.9964, Pperm = 0.009). CAP analyses revealed that the species composition of the timed swim dataset was the most consistently distinct with the highest classification success (100%), followed by the documentary video dataset and the timed stationary dataset (89% and 90% respectively) and last the timed transect dataset (87%, Fig. 3). The high classification success of the timed swim dataset is not surprising as the survey was purposely designed to exclude common bentho-pelagic species such as two-spot demoiselles (Chromis dispilus), Kermadec demoiselles (Chrysiptera rapanui) and blue maomao (Scorpis violacea). These bentho-pelagic species contributed strongly to the first axis of the CAP analysis, which separated the timed swim dataset at one extreme from the timed stationary dataset and documentary video dataset at the other extreme. A second a priori bias was the probable focus on charismatic ‘characters’ in the documentary video. Accordingly, fishes such as spotted black grouper (Ephinephelus daemelii) and bluefish (Girella cyanea) helped distinguish the documentary video dataset from the timed swim dataset. Nevertheless, even when the charismatic characters featured in the documentary video were removed from the analysis, the classification success of each survey method remained very high (> 80%, see Supplementary Fig. S4).
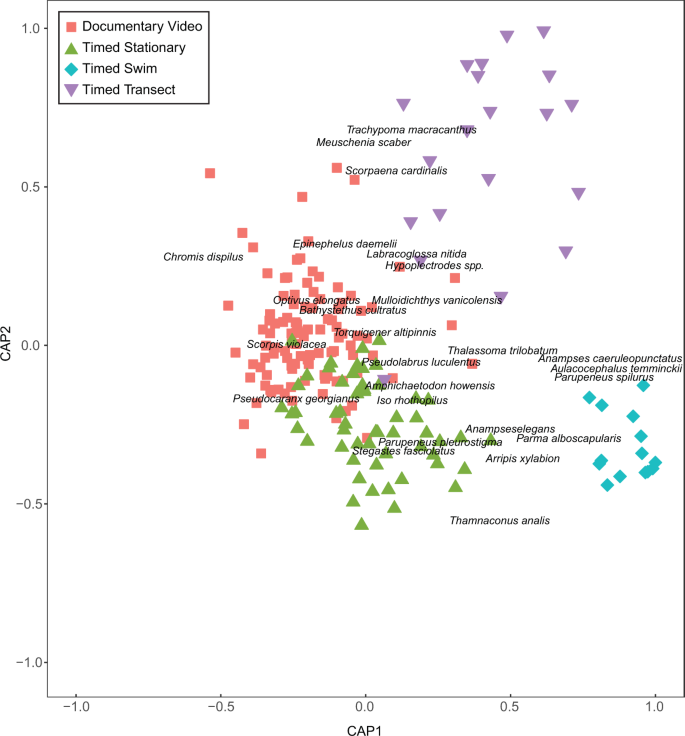
Surveyed fish community composition of Raoul Island, Rangitāhua. Shown are the first and second axes of the canonical analysis of principal coordinates for differences (expressed as Jaccards dissimilarity among samples) in fish community composition at Raoul Island, Rangitāhua for different survey methods. The ten most strongly correlated species (both positive and negative) for each axis are displayed to aid interpretation. Axes and sample coordinates have been scaled to unit length so that they are on the same scale as the correlations.
The timed transect, perhaps the most comprehensive and representative of the datasets, was separated from the other datasets on the second CAP axis, also suggesting a methodological difference. The species contributing strongly to the extremes of this axis were cryptic, stationary reef-dwelling species (such as Cook’s scorpionfish, Scorpaena cardinalis, and toadstool grouper, Trachypoma macracanthas) recorded in the timed transect and species potentially associated with the reef-sand interface recorded in the timed swim dataset (such as the darkvent leatherjacket, Thamnaconus analis and the blue-spotted wrasse, Anampses caeruleopunctatus) at the other extreme. These differences between the timed swim and timed transect are presumably driven by the survey design (as described in39). Specifically, the ‘timed transect’ was a belt transect, where the intention is to survey all fishes within a given area including cryptic fishes, whereas for the timed swim fishes were recorded to the edge of sight, increasing the potential for recording highly mobile fishes.
The disparate objectives of natural history film makers and research scientists will inevitably lead to shortcomings when re-purposing raw documentary video to infer biodiversity information. For example, the documentary video footage was not specifically created with the intention of taxonomically classifying all fish species, therefore not all fish could be identified. Furthermore, although rich metadata was provided in the shot log by the videographers (regarding subject matter, species, data, camera, videographer) this did not include geographic coordinates for the location of filming. Some location information was occasionally provided, and more specific details required cross-referencing the field logs and personal dive logs of the videographers. However, recording geographical coordinates in the shot log for future expeditions should be straightforward. Cameras and diving computers increasingly enable users to geo-reference events, and retrospectively retrieving coordinates using maritime charts or Google Earth is standard practice for many divers40.
Our survey of raw video taken for the purposes of creating a natural history documentary has provided valuable new biodiversity information for the remote, rarely surveyed, shallow reefs of Rangitāhua. Our study provides a proof-of-concept that raw documentary video has significant scientific value, and we demonstrate a method for strategically sampling raw footage to provide biodiversity information. Although the documentary video dataset found lower species richness than non-stationary methods and was biased toward large and charismatic fish species, traditional survey methods also recovered different species compositions from one another, depending on the survey design and objectives of each scientist. Overall, our study signals the untapped scientific value of thousands of hours of natural history video that exists throughout the world, which could be used to shine new light on rarely visited locations and retrospectively observe past baselines. There are often commercial restrictions on access to footage and publication of the data obtained from these sources that will need to be reconciled. Nonetheless, we anticipate that scientific inference from natural history video could increase in future as recording equipment able to capture high-resolution imagery becomes more accessible6 and advances in computer vision and machine-learning enables automated species recognition41,42.
Source: Ecology - nature.com
