Fungal status of the new winery before the arrival of the first harvest
The fungal diversity on the WREs of a new winery operating exclusively in spontaneous fermentation was studied to determine the fungal status of the winery before the arrival of the first harvest (T0 2016). Samples were collected from the floor, walls, new (NE) and used (UE) equipment. After Illumina sequencing, fungal OTUs were classified into 3 categories: (i) the mold and filamentous fungi genera, (ii) the yeast genera never described before in the winemaking process (named non-enological yeasts), and (iii) the yeast and yeast-like genera already described in the winemaking process (named wine-associated yeasts) (Fig. 1). Wine-associated yeasts include all the genera described in the literature in the vineyard, vine, grapes, must, wine and winery surfaces25,26,27,28,29.
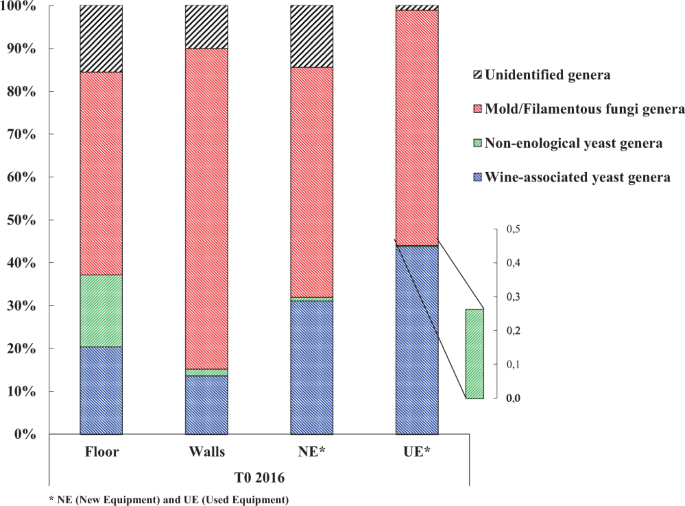
Percentages of the different fungal genera found on the different WREs of the new winery before the arrival of the first harvest (T0 2016).
Mold and filamentous fungi dominated the fungal genera identified on all the surfaces before the first harvest, with 294 different genera (Fig. 1 and Table 1). Yeast genera (non-enological and wine-associated) were identified before the arrival of any harvest in any of the WREs (Fig. 1 and Table 1) and a total of 35 different genera were found (Table 1). Wine-associated yeasts were identified on all the studied surfaces of the new winery and dominated the yeast population detected (Fig. 1 and Table 1). The identification of non-enological yeast genera may have resulted from the use of high-performance Illumina sequencing and the constant enrichment of the databases used7,30. The new winery surfaces (floor, walls and new equipment) showed a higher percentage (average 13%) of unidentified genera compared to the used equipment (1%) (Fig. 1).
Diversity and dynamics of mold and filamentous fungi genera
The different genera of mold and filamentous fungi identified at T0 2016 are represented using their relative abundances. Only the genera that represented >1% of relative abundance were selected (Fig. 2). Thus, a total of 32 mold and filamentous fungi genera among the 294 were identified on all the winery surfaces. At T0 2016, the taxonomic profiles observed on the floor, walls and the new equipment were quite close (with high representativeness of the genus Alternaria) compared to the used equipment which was dominated by the genus Cladosporium with the highest relative abundance (79%). These two genera had opposing relative abundances (a high abundance of Alternaria occurs with a low abundance of Cladosporium and vice-versa), suggesting potential interaction.
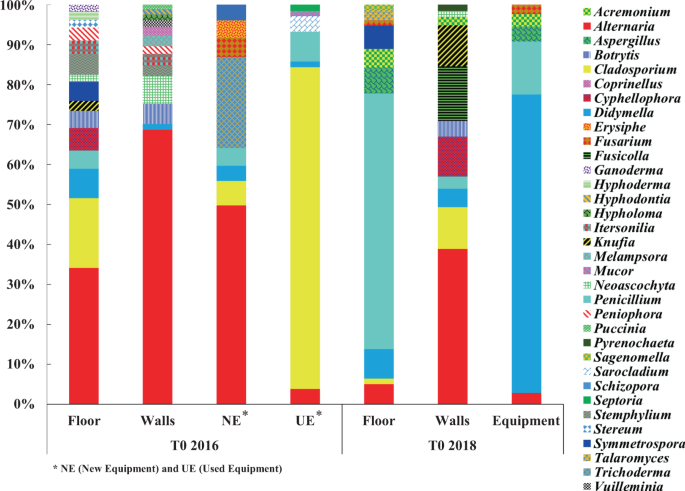
Relative abundance of the most represented mold and filamentous fungi genera identified on the different WREs prior to the 2016 and 2018 harvests. Represented genera are >1% relative abundance.
After two years of operations, a considerable modification of taxonomic profiles was observed, especially for the winery floor and equipment. These modifications resulted in a significant decrease in the total number of different identified genera (32 genera at T0 2016 and only 16 genera at T0 2018), as well as evolutions in the dominant genera (Fig. 2). So, the dominant genera Alternaria and Cladosporium described at T0 2016 were detected to a lesser extent at T0 2018, but persisted after 2 vintages. Their persistence throughout the whole study can be due to the presence of spores that might have been deposited from the surrounding atmosphere of the winery. Indeed, spores of Alternaria and Cladosporium are known to be widespread airborne spores31. Opposite abundances between the genera Alternaria and Cladosporium, observed at T0 2016, were also observed at T0 2018, suggesting again potential interaction (Fig. 2). After two years of operations, some genera became more widespread, like the genera Penicillium on the floor (57%) and Didymella on the equipment (68%). On the walls, Alternaria remained the most detected genus (35%), but new genera such as Cladosporium, Cyphellophora, Fusicolla, Knufia and Penicillium were also detected. The genus Acremonium was newly detected on all the WREs and the genus Aspergillus on the floor and equipment (Fig. 2).
Based on these results, implantation, colonization and persistence of some genera of mold and filamentous fungi were observed. For example: the genera Alternaria and Didymella present on all WREs between the vintages 2016 and 2018 can be considered as resident genera capable of persistence. After 2 years, genera Cladosporium and Penicillium are able to establish in all environments and persist. All these genera can therefore be qualified as ubiquitous genera. As for the genera Acremonium and Aspergillus, their implantation on environments might be linked to human activity and/or ventilation, which is an important factor of transfer of microorganisms between winery surfaces32. In addition, humidity and temperature described as two parameters influencing the growth of mold and filamentous fungi on surfaces like building material33,34,35 may also be factors influencing the establishment and persistence of the genera present on WREs.
Diversity and dynamics of non-enological yeast genera
Ten different genera of non-enological yeasts were detected before the arrival of the first harvest on all WREs. The genera Exophiala and Hannaella were the only genera present on all the winery surfaces with the highest percentage on winery floor (25% and 10%, respectively) (Table 2). After the 2 genera mentioned above, the genus Leucosporidium is the one with the highest percentages on winery floor (9%) and on walls (5%). These three genera are usually found in natural ecosystems like plants, soil, water and decaying wood material36,37,38,39. Therefore, it was not surprising to find these 3 genera on the surfaces of this new winery. These genera can therefore be qualified as ubiquitous genera.
Between T0 2016 and T0 2018, a significant increase in the biodiversity and quantity of non-enological yeasts were observed. Biodiversity-wise, 21 genera were detected at T0 2018 while only 8 genera were found at T0 2016. Quantity-wise, a 7-fold increase in the percentage of non-enological yeast was observed between these two time points on the walls (Table 2). Transfers of yeasts between different surfaces can be carried out by air flow and bioaerosol activity40 and over a period of one year27. Thus, these different transfers may explain the increase and dynamics of non-enological yeasts between T0 2016 and T0 2018. The two most abundant genera Exophiala and Hannaella, found initially at T0 2016, persisted in the winery environment throughout all the vintages and on all the winery surfaces. These two genera presented very high percentages, especially at T0 2017 on winery floor (73%) and T0 2018 on the winery walls (59%), however they did not exceed 5% on the winery equipment (Table 2). So, these results suggest that the conditions of every WRE plays an important role in retaining and shaping the fungal consortium of the winery. Indeed, the proliferation of fungi detected on different building material or in a particular area depends on the growth material and the conditions (e.g. humidity and temperature) under which they are found40,41. Compared to Exophiala and Hannaella, the genus Leucosporidium also persisted and colonized all WREs but with low percentages. Other genera previously detected at T0 2016, like, Buckleyzyma, Cutaneotrichosporon and Yamadazyma were detected frequently on all WREs but at very low levels (highest percentage: 2%) over the three vintages. Eleven new genera (e.g. Cystofilobasidium, Kondoa, Kregervanrija, Kwoniella, Lodderomyces, Malassezia, Oberwinklerozyma, Occultifur, Ogataea, Papilioterma, Piskurozyma, Pseudomicrostroma, Saitozyma, Schwanniomyces, Trichosporon and Udeniomyces) were detected punctually (1 or 2 times between T0 2016 and T0 2018) and the majority of them were detected after the fermentation activity (Table 2).
As for mold and filamentous fungi, different behaviors of non-enological yeasts were observed: some genera that can be considered as resident (environmental flora) and capable of persisting (Exophiala and Hannaella), genera capable of establishing and persisting (Buckleyzyma, Cutaneotrichosporon, Leucosporidium and Yamadazyma) and finally genera brought by the fermentation activity (e.g. Kregervanrija and Ogataea).
Diversity and dynamics of wine-associated yeast genera
At T0 2016, the taxonomic profiles for the floor, walls and the new equipment are quite close compared to the used equipment (Fig. 3), as described previously for mold and filamentous fungi. Relative abundance profiles show that the winery floor, walls and new equipment are primarily dominated by the yeast-like genus Aureobasidium (30%, 55% and 70%, respectively) and the genus Filobasidium (22%, 29% and 15%, respectively); however the used equipment is dominated by the genera Meyerozyma (45%), Wickerhamomyces (30%) and Aureobasidium (14%) (Fig. 3). Genus Aureobasidium was a dominant and common genus to all WREs, probably due to its capacity to persist in stress conditions linked to the sporulation ability of this genus42.
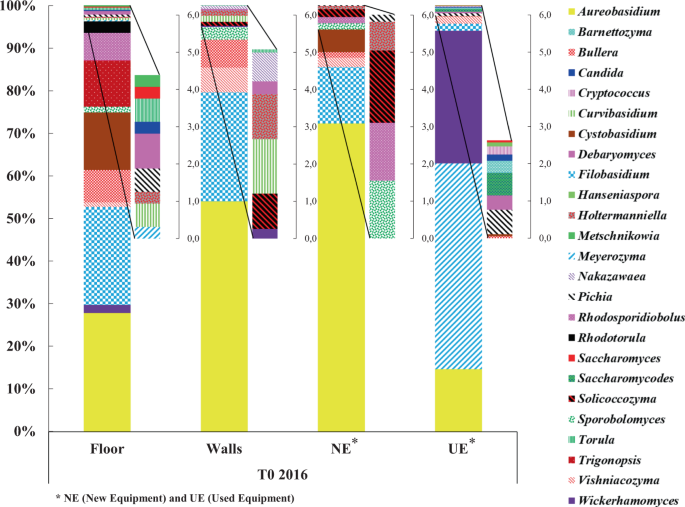
Relative abundance of wine-associated yeast genera identified on the different WREs of the new winery at T0 2016.
Before the arrival of the first harvest, yeast genera of enological interest like Candida, Hanseniaspora, Pichia and Saccharomyces were detected. Their presence on the used equipment is not surprising and is linked to the presence of must residues and/or wine, particularly in areas that are difficult to clean. The detection of the genera Candida, Pichia and Saccharomyces on the winery floor could be explained by the potential transfer between this environment and the used equipment due to human activities. Before the arrival of the first harvest, none of the yeast genera of enological interest were identified on the walls (Fig. 3).
Dynamics of wine-associated yeasts through three vintages on floor
On the winery floor at T0 2016, the 5 genera Aureobasidium, Candida, Cystobasidium, Filobasidium and Wickerhamomyces represented more than 65% of the total abundance (Fig. 4a). These genera were detected over time with the highest abundance: at T3 2016 (95%), at T0 2017 (92%), at T3 2017 (79%) and at T0 2018 (70%), but also with changes in their percentage of abundance. Indeed, a 75-fold increase of the genus Candida was observed at T3 2016, probably due to the fermentation activity (transfer from must and wine). The genera Bullera, Vishniacozyma and Metschnikowia detected at T0 2016 were also detected over time. The genus Saccharomyces detected at T0 2016 (<1%) was detected only at T3 2016 till T3 2017 but not at T0 2018. The genera Hanseniaspora, Priceomyces and Starmerella described on grapes and must19,42 were newly detected probably brought in during the harvest with the grapes and harvest equipment.
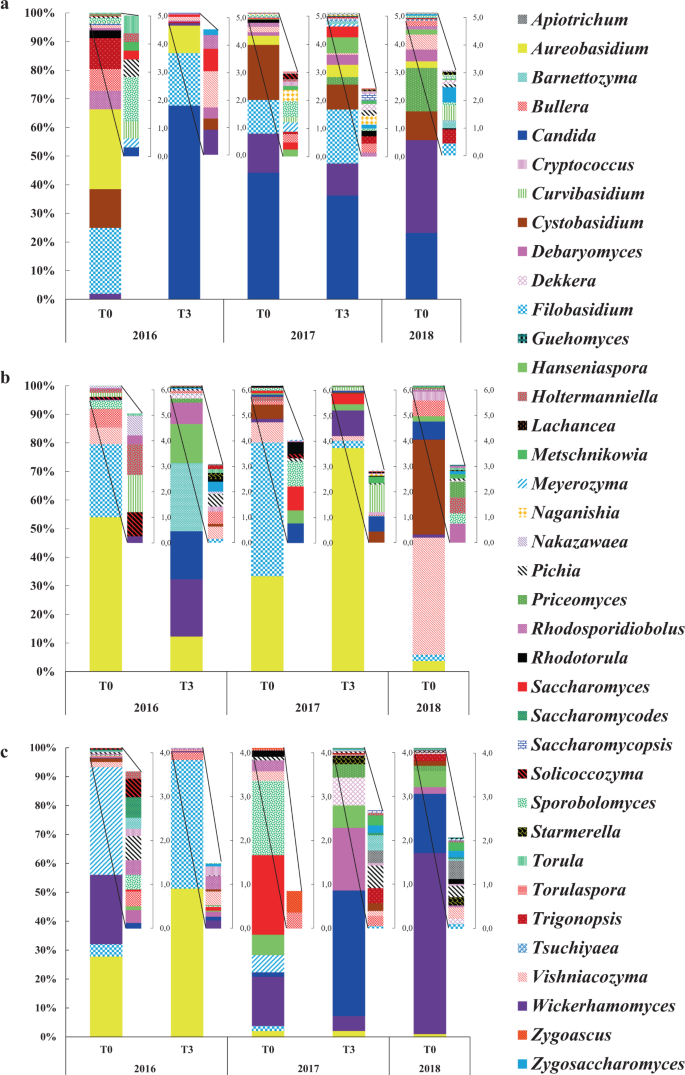
Relative abundance of wine-associated yeast genera identified on winery floor (a), walls (b) and equipment (c) throughout 2016, 2017 and 2018 vintages. Taxonomic profile at T0 2016 is the average of profiles obtained on old and new equipment.
During the 3 vintages, the presence of spoilage microorganisms like the genera Dekkera (as from T3 2017) and Zygosaccharomyces (as from T3 2016) was observed on the winery floor (Fig. 4a). The presence of these yeasts 3 months after the AF could be strongly linked to the fermentation activity (transfer from must and wine). However, their implantation and persistence took place on the winery floor only after the second harvest.
Therefore, we showed that the communities found at T0 2016 conditioned the future resident flora (T0 2018) on the winery floor by the presence over time of the same 16 genera, particularly the genera Aureobasidium, Candida, Cystobasidium, Filobasidium, Metschnikowia and Wickerhamomyces, while others (Solicoccozyma and Torula) showed punctual presence. We also showed that the AF might underlie the enrichment (Candida) or introduction of genera of interest (Hanseniaspora and Starmerella) and/or spoilage (Dekkera and Zygosaccharomyces) yeasts on the winery floor.
Dynamics of wine-associated yeasts through three vintages on walls
Some dominant genera on the walls were in common with those found on the floor, like Aureobasidium, Bullera, Filobasidium, Vishniacozyma and Wickerhamomyces. Their evolutions were similar as they were detected over the three vintages but changes in their percentage of abundance are observed. New genera like Candida, Cryptococcus, Cystobasidium, Hanseniaspora, Pichia, Saccharomyces and Starmerella were detected after the first or the second harvest (Fig. 4b). Their presence on the walls may have been due to transfer via grape berries and/or must because they are also commonly described on grape berries and must26,43 or to the result of transfers between WREs by air flow or bioaerosol activity. In addition, the increase in the abundance of some genera already present on the walls at T0 2016, such as Debaryomyces, may also have been due to fermentation activity and/or to the transfer of these genera between WREs. The spoilage genera Dekkera and Zygosaccharomyces were detected only at T3 2016 and at T0 2018 (Fig. 4b), and did not show real persistence.
Thus, after 2 years, the resident flora of winery walls included 6 wine-associated yeast genera (Aureobasidium, Bullera, Debaryomyces, Filobasidium, Vishniacozyma and Wickerhamomyces) found between T0 2016 and T0 2018, as previously described for winery floor. The AF activity also impacted the yeast flora of the walls (particularly the first fermentation) by the enrichment of certain genera like Debaryomyces and Wickerhamomyces and by the occurrence of new genera like Candida, Cryptococcus, Dekkera, Metschnikowia, Pichia, Saccharomyces, Starmerella and Zygosaccharomyces. The genus Saccharomyces was only detected during the 2017 vintage (T0 and T3), and did not persist in 2018. However, certain genera persisted until 2018 (e.g. Candida, Hanseniaspora). In addition, the flora of the walls showed radical changes at different time points. Thus, the resident flora of the walls appeared to have a higher state of instability than that of the winery floor. This instability may also be due to the fact that several genera, mainly of enological interest, were not in an environment favorable for their installation. Their presence appeared to be the result of transfers from other surfaces.
Dynamics of wine-associated yeasts through three vintages on equipment
At T0 2016 an average taxonomic profile was established by the association of the 2 profiles obtained from the used and new winery equipment (Fig. 4c). The genera Aureobasidium and Filobasidium, which were more abundant for the 2016 vintage (as for floor and walls), presented very low abundances afterwards (10% at T0 2017, 2% at T3 2017 and 1% at T0 2018). Throughout the vintages, the taxonomic profiles became enriched by other genera, most importantly, Candida, Debaryomyces, Hanseniaspora, Metschnikowia, Saccharomyces, Sporobolomyces and Wickerhamomyces which became more widespread (Fig. 4c). The presence of these genera, and more particularly for the genus Saccharomyces, probably resulted from must handling (beginning of AF in a fermentation tank) before the sampling of the winery equipment at T0 2017. After this time point, the genus Saccharomyces was not detected, so does not seem to persist. The higher presence of non-Saccharomyces yeasts compared to Saccharomyces on winery equipment have been previously reported44.
The spoilage genera Dekkera and Zygosaccharomyces were both probably provided by must and/or wine, particularly for the 2017 vintage. The highest abundance of the genus Dekkera was observed at T3 2017 and persisted to T0 2018 (Fig. 4c).
Thus, the changes of wine-associated yeasts on winery equipment showed specific dynamics compared to the floor and walls. Shifts in the taxonomic profiles were observed between the 2016 vintage and the next vintages. These observations and interpretations must take into account that winery equipment is regularly cleaned and is in direct contact with must and wine. Also, the results show that the evolution of wine-associated yeasts on equipment is not conditioned by the initial yeast population (T0 2016) as found for the winery floor and walls. Despite this, some wine-associated yeast genera appear to be ubiquitous and adapted to winery conditions. Indeed, the genera Aureobasidium, Bullera, Filobasidium and Vishniacozyma were detected throughout the whole study no matter which WRE it is.
Establishment of a winery consortium
The presence of mold, filamentous fungi and yeasts on the WREs of a new winery before the arrival of the first harvest was described. Considering the different abundances of all the genera (not only those which represent >1%), a total of 329 different fungal genera were identified at T0 2016 (294 mold and filamentous fungi and 35 yeasts) on all the WREs, which have never been described before. After 2 years, 172 fungal genera (132 mold and filamentous fungi and 40 yeasts) were identified and 17 fungal genera were newly detected (5 mold and filamentous fungi and 12 yeasts).
Among the total fungal genera and in terms of abundance, a total of 40 genera (mold, filamentous fungi and yeasts) presented different evolutions over time (Fig. 5). On the one hand, 10/40 genera (Alternaria, Aureobasidium, Bullera, Didymella, Exophiala, Filobasidium, Hannaella, Holtermanniella, Sporobolomyces and Vishniacozyma) were detected before the first harvest (T0 2016) and throughout the whole study on all the WREs until T0 2018 (Fig. 5). The majority of these 10 genera are commonly described on the berries and/or in must and have been considered, so far, as related to vineyard and wine-growing environments. However, according to the literature, these genera are described as ubiquitous genera widely distributed in natural ecosystems (soil, air, water and plants) and can colonize buildings45,46,47. In fact, the genus Alternaria is able to survive in cold, hot and dry weather and all year round in debris and seeds48,49,50. In addition, the sporulation capacity described for mold and fungi genera could contribute toward enhancing their persistence in adverse conditions like the winery environment34,51. Certain phenotypic traits described for the persistent genus Aureobasidium like the production of extracellular polysaccharides52 and film forming capacity53 may explain its persistence throughout the study. The persistence of Bullera, Filobasidium and Vishniacozyma genera may be related to their belonging to the Basidiomycetous yeast division and more particularly to the oxidative group of Basidiomycetous. Indeed, this group is described as being usually found in higher dominance in stressful environments and persisting well in adverse environments54,55. Several studies demonstrated the tolerance of the genus Exophiala to different stresses56 and the Hannaella yeast genus has been described in the phyllosphere of various plant species including Vitis vinifera and in soil35,57 which may explain their persistence over time. Thus, according to the results obtained in this study and the specific capacities of the genera described previously, we confirm that these 10 genera are not specific to a given environment like that of wineries. These genera are resident environmental flora that can persist in stressful environments including a winery (Fig. 5).
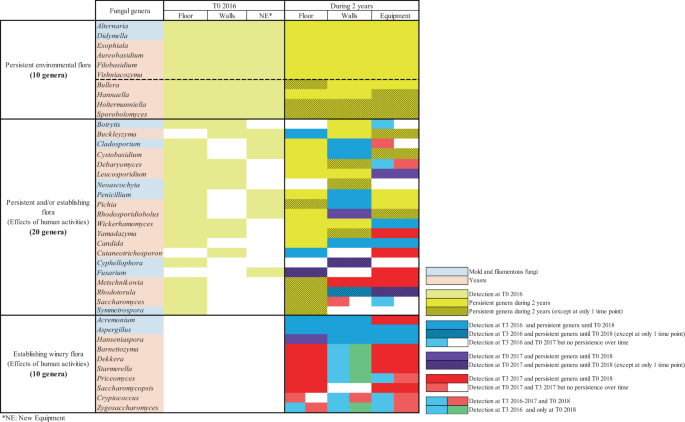
Resident and established fungal genera detected on the different WREs. Mold and filamentous fungi genera represented are >1% of relative abundance.
In addition, 20/40 of the fungal genera were detected at T0 2016 and showed persistence after 2 years on 1 or 2 WREs (Fig. 5). As previously described, ubiquitous character of these fungal genera linked to some phenotypic traits can explain their persistence37,39,54,55,58,59,60,61,62,63,64,65,66,67,68,69,70. These genera are considered as persistent and/or establishing winery flora. Among them, some genera became established via the harvest or transfers between WREs, especially on the walls and equipment (Fig. 5). After their implantation, these genera were able to persist until 2018, except the genus Saccharomyces, which persisted only on the floor 2 years after the arrival of the first harvest (Fig. 5). After the implantation linked to harvest, the persistence of the genus Candida may have been due to phenotypic traits like invasive growth71 and film formation53. Regarding the genus Pichia, the capacity of some species, isolated from grapes, to form ascospores72,73 and to produce killer toxins (proteins secreted by a fungi inhibiting the growth of other fungi)74 potentially contribute to the implantation and persistence of this genus in winery environment. Establishment and persistence of the genus Metschnikowia may be linked to its ability to outgrow other microorganisms due to the production of reddish pulcherrimin involved in iron chelation75,76. Furthermore, among the genus Wickerhamomyces, the species W. anomalus (previously known as Pichia anomala)77 is known for its resistance to osmotic pressure and the production of killer toxins78,79,80.
Finally, 10/40 of the fungal genera (Acremonium, Aspergillus, Barnettozyma, Cryptococcus, Dekkera, Hanseniaspora, Priceomyces, Saccharomycopsis, Starmerella and Zygosaccharomyces) were not detected at T0 2016 but exhibited implantation after the 2016 and/or 2017 harvests and certain of them persisted over time depending on the WREs (Fig. 5). The capacity of the genus Aspergillus to produce conidia81 and the antagonism activity of the genus Acremonium82 can explain their establishment in winery environment. Regarding the genus Hanseniaspora, the implantation and the persistence of 2 species of Hanseniaspora during 2 vintages has been previously demonstrated19. The genus Priceomyces has never been described in a winery environment before, only on grape berries43. Nevertheless, this genus was detected over one year in brewery air83. Thus, all yeast genera showed potential for implantation in all WREs and to become part of the established winery flora as well as the spoilage genus Dekkera previously described as a persistent genus in several steps of the winemaking process and in WREs84,85. So, the implantation of all 10 genera can be explained by human activities (fermentation and/or transfer) in the winery and are considered as establishing winery flora (Fig. 5).
In conclusion, this work provided new insights into the establishment of the fungal consortium in a new winery. This fungal consortium consisted of persistent environmental flora that is not specific to the winery environment. These environmental flora included mold, filamentous fungi (e.g. Alternaria, Didymella) and yeasts (e.g. Aureobasidium, Filobasidium). Certain genera (e.g. Candida) were found before the arrival of the first harvest on one WREs, mainly on the winery floor, and persisted over time. Meanwhile, on the other WREs, the same genera were detected following the start of the harvest. Therefore, this finding suggests that these genera are well adapted to one specific environment of the winery and could be classified as persistent flora and/or establishing flora. However, during 2 vintages, the environmental flora was enriched by other fungal genera (e.g. Acremonium, Hanseniaspora). These genera were considered as establishing winery flora and are less adapted to the winery environment. Moreover, the majority of establishing flora are genera that present a fermentative metabolism that could also to be a prerequisite state preventing their adaptation. Future works could be carried out to understand the mechanisms implicated in the establishment and persistence of this flora on the WRE of this new winery and more particularly, flora of enological interest.
Source: Ecology - nature.com
