Colony collection and maintenance
Three colonies of M. graminicola were collected from natural nests (inside oak galls fallen on the ground) located in a leaf litter of a wooden area in Fornoli (MS) (Tuscany, Italy). Colony fragments (about 40–50 workers, a queen and some brood each) together with their original gall nests were housed in plastic arenas (10 × 10 × 3 cm) with a plaster floor on which a 2 cm layer of sifted organic material taken from the field was placed. Laboratory conditions were controlled (T 20°/23 °C, RH 50/60%) with a 12 h light-dark cycle. Colonies were fed on a mixed solution of water, sugar and mealworms fragments. Small pieces of pumpkin seeds were also provided.
Experimental set-up
For each trial of each experiment 60 replicates (except for experiment 3 and 6, see below) were conducted and single ants (20 from each colony) were tested only once for each treatment. Trials were carried out randomly from 10.00 a.m. to 4 p.m. To study ant reactions to a potential danger (a simulation of predator/enemy approaching or other distress situations causing ground vibrations or loss of contact with the substrate) we offered different kind of stimuli as described below.
Reactions to disturbance on horizontal plane (Experiment 1)
The ants were introduced into experimental arenas (10 × 10 cm, floor covered by filter paper) and after 5 s, their possible reactions (freezing, walking, body curling into a ball) to the following stimuli were recorded:
- (1)
Slight tapping of the horizontal plane by a plastic stick (n = 60).
- (2)
Strong tapping of the horizontal plane by a plastic stick making the ant bounce (n = 60).
- (3)
Slight tapping on the ant gaster by the tip of an entomological forceps (n = 60).
- (4)
Strong tapping on the gaster by the tip of an entomological forceps causing the ant to lose contact with the substrate (n = 60).
Walking ability on inclined planes and reaction to disturbance (Experiment 2)
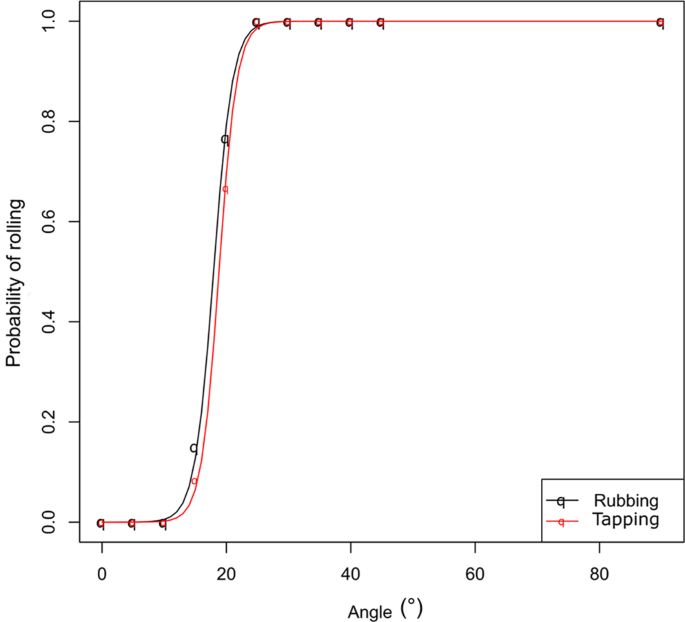
The effect of slopes on the walking ability of the ants was tested allowing single individuals to freely move on inclinable plastic plane (5 × 17 cm) covered by filter paper (changed after every trial) with a 3 cm horizontal platform on which the ants were released. The number of ants that covered the entire path length and those eventually falling down (within 5 min experimental time) were recorded. Different inclinations of the plane were tested, from horizontal (0°) to 45° (every 5°). Vertical inclination (90°) was also tested (n = 60 for each inclination).
This was preliminary to a second set of experiments aimed to test how the ants behave upon receiving mechanical stimuli without being directly touched while moving along inclined planes (experimental device and gradients as above). As stimuli, we offered different slight vibrations on the ground that did not cause the ants losing the contact with the substrate:
- (a)
Single slight tap with a plastic stick at the base of the experimental device (n = 60).
- (b)
Slight substrate rubbing using an entomological forceps just behind the walking ant (avoiding any contacts with the ant body) (n = 60) (Supplementary Video S1).
In both cases, stimuli were offered ~5 sec after the ant release.
Analysis of different phases of body curling and active rolling on inclined plane (Experiment 3)
Using the same conditions and experimental device as described in Exp. 2b, we recorded and analysed 10 videos (each involving a different individual) for a fine analysis of ant movements. Here and for all further experiments on inclined planes, a gradient of 25° was chosen because this was the critical slope for a 100% probability of rolling by the ants (see Fig. 1) (n = 10). For a better understanding and description of the different movements during rolling, we made a frame-by-frame analysis on macro videos. This allowed a better visualization of the sequence of movements of the body parts. Then we played the videos at slow motion (25% reduced speed), in order to observe and better describe the whole process.
Recordings were performed at 60 fps (progressive) using Canon 80D and 24 fps (progressive) using Blackmagic Cinema Camera Pocket with Metabones Speedbooster Adapter; Lens: Canon 100 mm Macro f/2, 8 IF USM, Sigma 24-105 f/4 DG OS HSM, Tamron 85 f/1, 8 Di VC USD; Tripods: Manfrotto 055X Pro with head Manfrotto 808 RC4, Manfrotto MK290 Dual with head 128RC and Edelkrone Slider One. To illuminate the experimental area, a set of three cold light lamps (85 W) was used with soft box (60 × 60 cm). For video editing and digital crop Final Cut Pro X software was used.
The clips (60 fps progressive) were analysed frame-by-frame by Kinovea software (www.kinovea.org) and a sequence of movements was described as a general pattern under those specific experimental conditions. The “reaction time” was measured as the time from the frame at which the forceps touched the substrate to the frame at which the antennae started moving backwards to prepare rolling. The “latency to roll” was measured as the time from the frame at which the forceps touched the substrate to the frame at which the ant started rolling downwards.
To measure the linear speed (distance/time) of the rolling ants (approximated to points), we analysed 20 videos each involving a different ant. Rolling was induced as in Exp. 2b. However, in this case to evaluate the distance covered from the starting point to the end of the rolling path the plane was covered by a graph paper. The duration of the movement was measured by Kinovea software (n = 20). By the same methods, we also evaluated the walking speed of the ants (n = 12) just before being offered the stimulus that induced rolling.
Duration of the ball-like position (Experiment 4)
We measured how long the ants (singly tested in the same arena as in Experiment 1) kept the ball-like position upon receiving three different strong distress stimulations. (A) Tapping the ants from behind with the tips of a forceps making them loosing contact with the substrate; (B) vertical drop from a height of 10 cm (the ants were slightly pushed down from a horizontal plane); (C) active rolling on a 25° inclined plane after substrate rubbing with forceps (as in Exp. 2b) (n = 60 for each treatment).
Rolling on natural substrates (Experiment 5)
In this set of experiments, the rolling behaviour on natural substrates (leaves, earth and stones) was evaluated using materials collected from the field area where colonies were found. Leaves, earth or stones were placed on an inclined plane (25°) on a wooden support (15 × 7 × 13 cm). The ants were introduced on the device and singly tested. Disturbance was obtained by substrate rubbing with an entomological forceps just behind the walking ant as in Exp. 2b. For each trial, we recorded if the ant rolled down and the distance reached before resuming a normal walking or before going out of sight (vanishing point) in the case of leaves and stones (n = 60 for each treatment).
Reaction to aggression (Experiment 6)
Reaction to aggression was tested and the effectiveness of rolling as a defensive strategy evaluated. Aggression tests with other ants (Temnothorax unifasciatus collected in the same area of M. graminicola), were carried out in fighting boxes (4 × 4 × 2 cm, floor covered by filter paper) where dyads were introduced. Tests (experimental time 10 min) were conducted on both horizontal and inclined planes (25°, 20 cm long). A paper bulkhead separating the ants was removed 5 min after the ants were introduced into the box. In experiments on inclined plane, a second bulkhead (representing one of the wall of the box) was removed allowing the ants eventually to roll down to a second arena (10 × 10 × 2 cm). For this experiment, 30 replicates (10 ants from each colony) of each kind of aggression test (horizontal/inclined) were carried out. Ants were tested only once for each treatment.
A general description of the ants’ behaviour and the following occurrences were recorded:
- (a)
Ant initiating the attack;
- (b)
Curling of the body into a ball;
- (c)
Rolling behaviour (only for inclined plane);
- (d)
Defensive effectiveness of rolling (only for inclined plane), measured as the percentage of Myrmecina ants disentangling from the Temnothorax attack during or at the end of the rolling;
- (e)
Number of injured or dead ants;
- (f)
Attack time (total time spent on aggression).
Statistical analysis
Chi-square test (2 × 3 tab) and following Analysis of Standardized Residuals were used to evaluate data from Exp. 1 (software: IBM SPSS Statistics, Italian version 24). A Logistic Regression Model was used for analysis of data from Exp. 2 (software: R version 3.3.1, www.R-project.org). Kruskal-Wallis test followed by pairwise multiple comparisons (Bonferroni correction) were used for analysis of data from Exp. 4 and Exp. 5, Mann-Whitney U test and Chi square test for Exp. 6 (software: IBM SPSS Statistics, Italian version 24).
Throughout the text, average values are reported with ±standard errors, while median values (mdn) are reported with minimum-maximum (min-max) range values.
Animal use and welfare
All experiments reported in the present research were performed in accordance with relevant guidelines and regulations on animal use.
Source: Ecology - nature.com
