Different amounts of silk building materials
A single H. euryalus cocoon from our sample contained only an outer envelope, i.e., no inner envelope was spun, but the larval pellet and pupal casing were found inside this otherwise normal (size and shape) outer envelope. This cocoon was therefore omitted from our silk analyses.
We found that the cocoons from the different Hyalophora groups contained different amounts of total cocoon silk (One-way ANOVA: F5, 58 = 18.7600, p < 0.0001). Post hoc comparisons (Tukey HSD test, ∝ = 0.05) showed that loose/round (H. euryalus) and baggy (H. cecropia) cocoons had a higher amount of silk than cocoons with compact morphology (p < 0.0001 for all comparisons between H. euryalus and compact cocoon groups, and for all comparisons between baggy H. cecropia and all compact cocoon groups). Hyalophora euryalus and H. cecropia, however, were indistinguishable from each other (H. euryalus: 0.8726 +/− 0.0537 g, n = 9; H. cecropia: 0.7800 +/ 0.0389 g, n = 10). Between the two H. cecropia cocoon morphologies, compact cocoons (0.6800 +/ 0.0389 g, n = 10) had similar amounts of total silk as baggy cocoons, but contained significantly less silk than H. euryalus cocoons. For the three species with monomorphic compact cocoons, the cocoons of H. cf gloveri (0.6292 +/ 0.0486 g, n = 10) had intermediate amounts of total silk, and the cocoons with the least amounts of total silk belonged to H. “kasloensis” (0.4851 +/ 0.0403 g, n = 10) and H. columbia (0.3786 +/ 0.0293, n = 10). Hyalophora groups that produce polymorphic cocoons (H. cecropia and H. euryalus) contain more total silk than groups that produce monomorphic cocoons (p < 0.0001 for all post hoc comparisons between each of H. cecropia and H. euryalus, with that of H. cf gloveri, H. “kasloensis”, and H. columbia, respectively).
We found that cocoons from the different Hyalophora groups significantly differed in their percentage of total cocoon silk that was partitioned to the outer envelope (Kruskal-Wallis test: χ2 (5) = 45.3088, p < 0.0001). Post hoc Wilcoxon pairwise comparisons (∝ = 0.05) found that although loose and round H. euryalus cocoons had a significant percentage of total silk in the outer envelope, overall, groups that spun compact cocoons allocated the greatest proportion of total silk to the outer envelope. Compact H. “kasloensis” cocoons were similar to H. euryalus cocoons, and also had the greatest percentage of total cocoon silk in the outer envelope. Compact H. columbia and compact H. cf gloveri cocoons had intermediate percentages of total silk devoted to the outer envelope, followed by H. cecropia compact cocoons. Baggy H. cecropia cocoons had the lowest percentage of total silk devoted to the outer envelope.
Cocoons from the different Hyalophora groups significantly differed in their percentage of total cocoon silk found in the intermediate space of cocoons (Kruskal-Wallis test: χ2 (5) = 48.5039, p < 0.0001). Post hoc Wilcoxon pairwise comparisons (∝ = 0.05) showed that baggy H. cecropia cocoons had the greatest percentage of total silk in the intermediate space, and this percentage was significantly greater than the percentages of all other groups. Compact H. cecropia cocoons had the next largest percentage of total silk in the intermediate space, followed by H. euryalus, H. cf gloveri, and H. “kasloensis” cocoons, which all had similar percentages of total silk. Hyalophora columbia cocoons contained no silk in the intermediate space.
The cocoons from the different Hyalophora groups differed in the percentage of total silk found in the inner envelope (Kruskal-Wallis test: χ2 (5) = 37.3455, p < 0.0001). This difference was not as marked across groups, however, as this difference was due to a significant difference between two clusters: baggy H. cecropia, compact H. cecropia, H. columbia, and H. cf gloveri having a significantly greater percentage of total silk in the inner envelope than H. euryalus and H. “kasloensis” (post hoc Wilcoxon pairwise comparisons, ∝ = 0.05).
Outer envelopes – size and shape
No significant interaction between Hyalophora group and total cocoon silk (ANCOVA: F5, 5) = 0.5788, p = 0.7159) was found in our initial comparison of outer envelope surface areas. In our subsequent ANCOVA analysis with the interaction term removed, we found that the surface areas of cocoons were significantly different between the Hyalophora groups (F(5, 5) = 75.7264, p < 0.0001; Fig. 3a). Post hoc comparisons (Tukey HSD test, ∝ = 0.05) found that baggy H. cecropia cocoons had the greatest outer envelope surface areas, followed by compact H. cecropia and H. euryalus cocoons; all of which spin polymorphic cocoons (Fig. 3a). The cocoons with the lowest surface areas were H. “kasloensis”, H. cf gloveri, and H. columbia. Cocoons from these three species were similar in surface area and all produce monomorphic cocoons (Fig. 3a). A positive relationship between total cocoon silk and outer envelope surface area was also found (F(1, 1) = 5.0187, p = 0.0294).
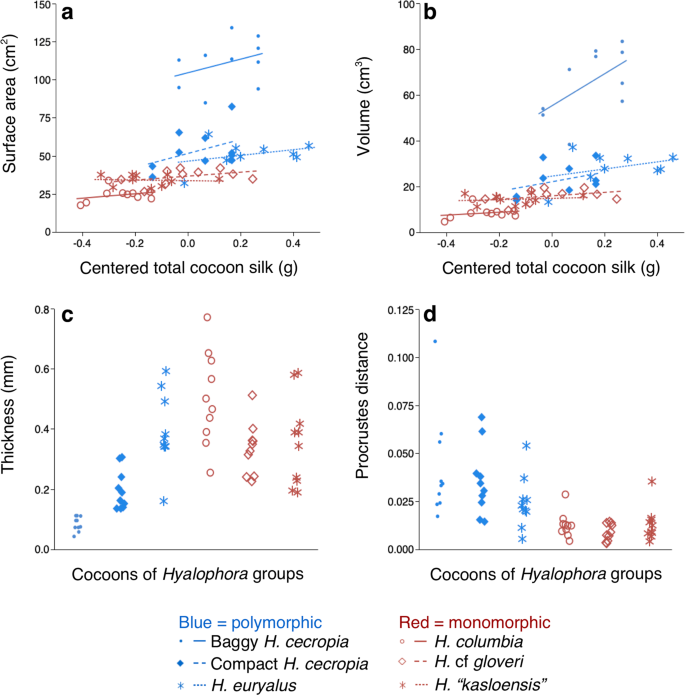
Comparisons of the architectural features of the outer envelopes of cocoons from the different Hyalophora groups that we examined. (a) Surface area. (b) Volume. (c) Thickness. (d) Shape (as measured using Procrustes distance22,23). For (a–d), data were analyzed using ANCOVA (n = 10 for all groups).
Similarly, we found no significant interaction between Hyalophora group and total cocoon silk (ANCOVA: F(5, 5) = 1.9169, p = 0.1093) when we initially compared outer envelope volumes. In our subsequent analysis with the interaction removed, we found that the outer envelopes differed in volume (F(5, 5) = 54.8507, p < 0.0001; Fig. 3b). Baggy H. cecropia cocoons had outer envelopes with the greatest volume relative to all other cocoons (post hoc Tukey HSD test, ∝ = 0.05; Fig. 3b). Cocoons from all of the other Hyalophora groups had similar outer envelope volumes (Fig. 3b), and a positive relationship between total cocoon silk and outer envelope volume was observed (F(1, 1) = 7.2323, p = 0.0096).
In comparing outer envelope thicknesses, we also found no significant interaction between Hyalophora group and total cocoon silk (ANCOVA: F(5, 5) = 1.0996, p = 0.3733). Without the interaction term in our analysis, we found that cocoons from the different groups differed in outer envelope thickness (F(5, 5) = 16.5639, p < 0.0001; Fig. 3c), but thickness was not related to the type of cocoon spun (Post hoc Tukey HSD test, ∝ = 0.05; Fig. 3c), nor was thickness related to total cocoon silk (F(1, 1) = 2.6057, p = 0.1125).
In our initial comparison of outer envelope shape, there was no significant interaction between total cocoon silk and Hyalophora group (ANCOVA: F(5, 5) = 0.7988, p = 0.5561). In our succeeding analysis with the interaction term omitted, we found that the outer envelopes differed in shape (F(5, 5) = 5.5116, p = 0.0004; Fig. 3d) between groups. The outer envelopes segregated into the following groups according to overall shared shape: H. euryalus and H. columbia cocoons had similar shape; the rest of the Hyalophora groups each had outer envelopes with their own, different shape (post hoc Tukey HSD test, ∝ = 0.05). No relationship between total cocoon silk and outer envelope shape was found (F(1, 1) = 0.2110, p = 0.6479).
Intermediate space volumes
We omitted from our analysis a single H. euryalus cocoon that had no inner envelope. For one H. columbia cocoon, the intermediate space volume could not be measured, as the two distinct envelope layers were spun so close together; this cocoon received a value of zero for intermediate space volume. We found that the intermediate space volume was significantly different between the Hyalophora groups (Kruskal-Wallis test: χ2 (5) = 47.8515, p < 0.0001; Fig. 4). Cocoons from groups that can spin polymorphic cocoons had the largest intermediate spaces: baggy H. cecropia cocoons had the largest intermediate spaces of all cocoons; H. euryalus and compact H. cecropia cocoons were similar, and had the next largest intermediate space volumes (post hoc Wilcoxon pairwise comparisons, ∝ = 0.05). In contrast, the groups that spin monomorphic cocoons had the smallest intermediate spaces (Fig. 4). Hyalophora “kasloensis” and H. cf gloveri cocoons had similar, low intermediate space volumes, while H. columbia cocoons had the smallest intermediate space volumes of all groups.
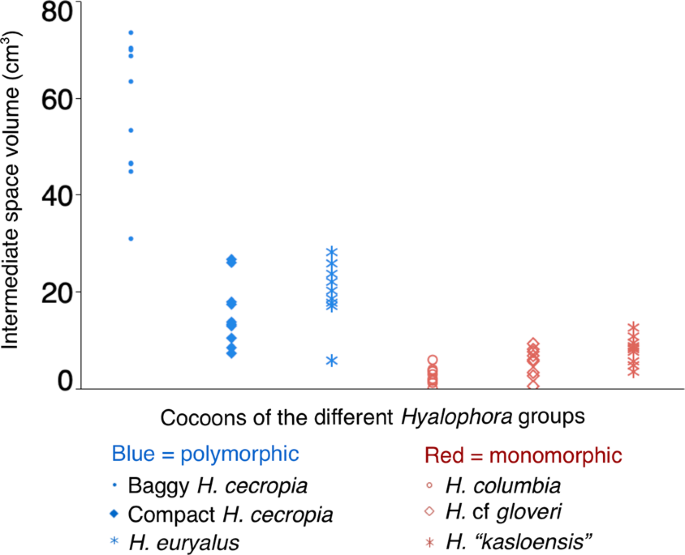
Comparison of the intermediate space volume of cocoons from the different Hyalophora groups observed in our study. Data were analyzed using the Kruskal-Wallis test (n = 10 for all groups, except for H. euryalus in which n = 9).
Inner envelopes – size and shape
In these comparisons, we omitted the H. euraylus cocoon that was without an inner envelope. We also omitted the H. columbia cocoon of which it was impossible to separate the inner envelope from the outer envelope properly. Here, the two distinct envelope layers were spun so close together that removal of the inner envelope resulted in an inner envelope layer that was too torn for 3D scanning.
No interaction between Hyalophora group and total cocoon silk was observed when we compared the inner envelopes of the Hyalophora groups in both surface area (ANCOVA: F(5, 5) = 0.5817, p = 0.7138) and volume (ANCOVA: F(5, 5) = 0.6839, p = 0.6379). Our subsequent analyses with the interaction term removed showed that the inner envelopes were different among the Hyalophora groups in both surface area (F(5, 5) = 11.4633, p < 0.0001; Fig. 5a) and volume (F(5, 5) = 13.6788, p < 0.0001; Fig. 5b). Hyalophora c. gloveri inner envelopes had both the greatest surface area and volume of all groups, and all other groups had similar inner envelope surface areas and volumes (post hoc Tukey HSD test, ∝ = 0.05; Fig. 5a-b). Total cocoon silk had a positive relationship with both inner envelope surface area (F(1, 1) = 28.6158, p < 0.0001) and volume (F(1, 1) = 37.2155, p < 0.0001).
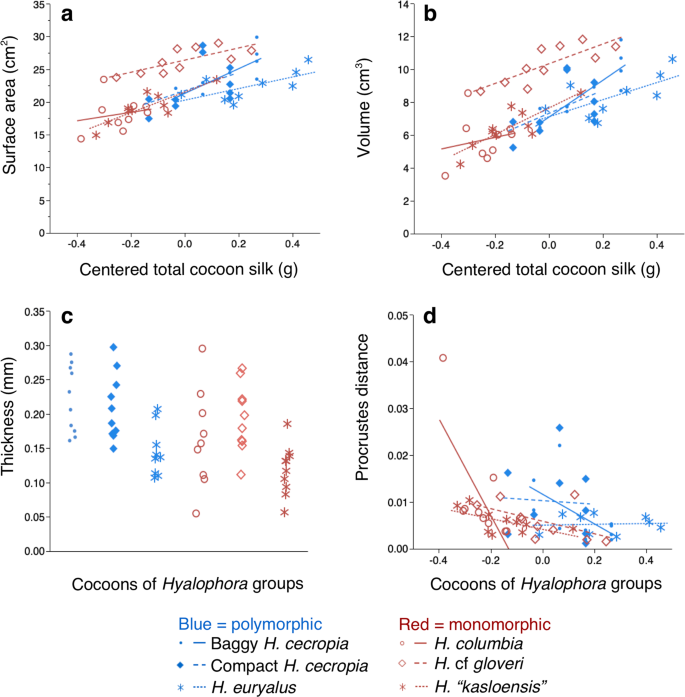
Comparison of the architectural features of the inner envelopes of cocoons from the different Hyalophora groups that we examined. (a) Surface area. (b) Volume. (c) Thickness. (d) Shape (as measured using Procrustes distance22,23). For (a–d), data were analyzed using ANCOVA (n = 10 for all groups, except for H. euryalus and H. columbia in which n = 9).
An initial comparison of inner envelope thicknesses found no interaction between Hyalophora group and total cocoon silk (ANCOVA: F(5, 5) = 1.7775, p = 0.1363). We found that the inner envelopes differed in thickness (F(5, 5) = 7.7078, p < 0.0001; Fig. 5c) in our subsequent analysis that omitted the interaction term, but inner envelope thickness was not related to either cocoon-type (baggy or compact) or the number of cocoon-types that could be spun (polymorphic or monomorphic) (post hoc Tukey HSD test, ∝ = 0.05; Fig. 5c).
When we compared the shapes of the inner envelopes of the different Hyalophora groups, we found a significant interaction between Hyalophora group and total cocoon silk (ANCOVA: F(5, 5) = 3.2718, p = 0.0131; Fig. 5d). This result shows that the shapes of the inner envelopes of the Hyalophora groups will be different for inner envelopes at different sizes.
Evolution of cocoon morphology
The species tree generated by combining available COI datasets within this group, and removing all proposed hybrids or misidentified individuals, matches previous reconstructions for this clade12,13. In particular, this species tree highlights a clear divergence from the outgroup followed by a potential burst of species in Hyalophora lineages (characterized by low posterior probabilities at each of the nodes and short branch lengths; Fig. 6). The limitations of the small COI dataset, and the apparent rapid diversification within Hyalophora, put the exact character evolution of cocoons within Hyalophora beyond the scope of the current work, however. Nevertheless, our analysis demonstrates that Hyalophora has diverged from its outgroup (C. promethea) as cocoon construction in Hyalophora has evolved to include the production of separable, distinct envelopes, and the loss of a silk leaf (which attaches the cocoon to a branch). These traits, and the addition of silk to the intermediate space between envelopes in some of the Hyalophora lineages, are not found in the sister clade. Although little is known about cocoons of the newly described H. mexicana, and H. leonis lineages, the existence of cocoon polymorphism in the outgroup as well as in multiple ingroup species, suggests that cocoon polymorphism is an ancestral state. All Hyalophora monomorphic species constructed cocoons with compact morphology, though the uncertainty in phylogenetic relationships limits the interpretation of whether or not this has resulted from a single or multiple loses of polymorphic cocoon shape.
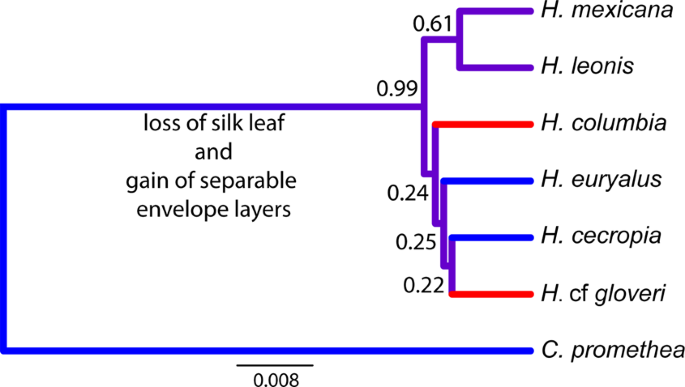
Phylogenetic species tree of Hyalophora silk moth species with C. promethea outgroup. This tree represents individuals from previous published trees12,13, with all hybrid and potentially misidentified individuals removed. Posterior probabilities are indicated at each node, highlighting uncertainty of branching order amongst the rapidly diverging Hyalophora lineages. Blue branches indicate species constructing polymorphic cocoons and red branches indicate species that construct monomorphic, compact cocoons only. Purple branches indicate that the presence or absence of polymorphism is unknown (H. mexicana and H. leonis).
Source: Ecology - nature.com
