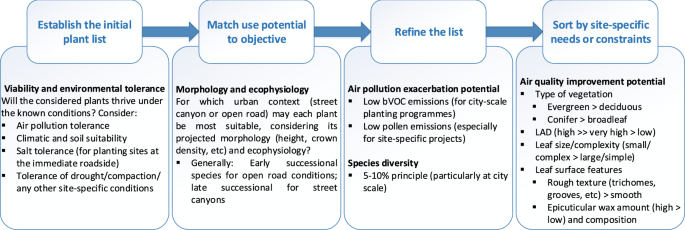
The diverse influences of individual species on air quality are reflected by the immeasurable range of phenotypes. As well as consideration of the planting environment, effective plant selection for air pollution mitigation requires an understanding of the balance between beneficial and detrimental aspects of vegetation at species level and below. In the following sub-sections, we critically evaluate major findings regarding the cost-benefit nature of species selection for urban air quality management.
Disadvantageous aspects of GI for air quality
Beyond potential interruptions to atmospheric dispersion within the urban boundary layer56, vegetation can be a source of additional air pollutants, primarily in the form of biogenic volatile organic compounds (bVOCs) and pollen. Indeed, the negative impacts upon air quality associated with such emissions may counteract or even outweigh any intended pollution abatement40. In order to minimise any disadvantageous aspects, careful and contextual species selection is critical42,57.
Biogenic volatile organic compounds
bVOC emissions from vegetation constitute a substantial global source of reactive hydrocarbon gases, including, most notably, isoprene, monoterpenes, and higher terpenoids58,59. bVOCs undergo atmospheric oxidation that, when combined with NOx, forms ground-level ozone (O3), a noxious gas with acute and chronic impacts on human respiratory and cardiovascular systems60. Secondary organic aerosols can also be produced on oxidation, potentially increasing ambient PM concentrations, whose negative impacts on human health are well-supported1,61. bVOC emissions are generally low below 20 °C, but increase with temperature until peaking between 35 °C and 40 °C (plant species-specific), at which point emission rates stabilise and begin to diminish42. The incidence of bVOC emissions in the United Kingdom may therefore increase over the coming decades under projected climate change scenarios59,62.
High bVOC emissions are particularly problematic in urban areas (e.g. Ren et al.63), where elevated temperatures and anthropogenic emissions converge38. bVOC emissions are also increased by other typical urban environmental stressors, including plant tissue damage by, for example, management interventions38. Plant tissue damage may instigate additional emissions in the form of green leaf volatiles (C6 aldehydes and ketones), implying the need to minimise pruning or other ‘wounding events’ (see ‘Sustainability and species suitability’). Moreover, a recent review by Fitzky et al.59 suggests the potential for positive feedback in the form of plant tissue damage by O3. The restriction of overall bVOC emissions from urban GI is therefore crucial38, and may be achieved by judicious plant selection.
Given that the atmospheric reactivity of many bVOCs is higher than that of comparable carbon number anthropogenic VOCs, it is important to consider not only the total amount but the individual type of bVOCs emitted42,64. The emission of bVOCs that exhibit higher reactivity should be avoided first and foremost, particularly in near-road environments. For example, some sesquiterpenes can have a lifetime of just a few minutes, enabling the generation of secondary aerosols in the ambient air even where dispersion from the source is unhindered42. However, there is limited available information on the causes, means and rates of production of specific bVOCs by individual plant species59. Relevant studies have tended to focus on the two most abundant bVOCs (isoprene and monoterpenes; e.g. Benjamin and Winer65), yet up to a hundred different bVOC species bear considerable influences in the atmosphere66. A comprehensive review by Laothawornkitkul et al.66 investigated the role of bVOCs at the global scale and found that anthropogenic environmental changes, including climate change, may perturb the natural relationship between the biosphere and the atmosphere, resulting in an overall increase in future bVOC fluxes. Primary sources of isoprene and monoterpenes include several genera of common UK urban vegetation, such as Populus, Salix and Platanus (for isoprene), and Quercus, Malus and Pinus (for monoterpenes)66. Other reactive bVOCs and less reactive bVOCs (with atmospheric lifetimes of <1 day and >1 day, respectively) were grouped and categorised as such, and major emission sources of both were identified as grassland as well as typical urban vegetation genera, including Betula66.
Beyond genus, bVOC emissions can vary by several orders of magnitude between species58. In a UK-based study, Donovan et al.40 developed an Urban Tree Air Quality Score (UTAQS) that ranks trees according to their potential for pollutant deposition versus bVOC emissions, based upon different modelled tree coverage and temperature scenarios. Resultant changes in O3 under each scenario were used to indicate that high bVOC emissions by some species lead to increased (0.8–2.9%) regional O3, whereas other species facilitated a modest decrease (0.3–0.8%) in O3 at the regional scale, due to a minimal increase in bVOC emissions relative to increased deposition40. Of the 30 tree species included in the study, 7 were found to have a ‘low’ UTAQS, defined as resulting in worsened air quality compared to the control scenario via increased concentrations (<3%) of O3 and NO2 despite enhanced HNO3 deposition: Populus tremula, Quercus petraea, Quercus robur, Quercus rubra, Salix alba, Salix caprea and Salix fragilis40. These seven species are considered to be high isoprene emitters. The highest-scoring (i.e. most beneficial) species were found to be: Acer campestre, Acer platanoides, Alnus glutinosa, Betula pendula, Chamaecyparis lawsoniana, Crataegus monogyna, Larix decidua, Prunus laurocerasus and Pinus nigra40. These nine high-scoring species are low isoprene emitters, but it should be noted that some (e.g. Pinus nigra) can release significant amounts of monoterpenes40,66.
As average UK temperatures increase and while anthropogenic emissions remain high, the development of comprehensive bVOC emissions profiles for viable GI species is recommended66. However, the strict classification of plant species as positive or negative according to bVOC emissions is highly problematic67, due in part to intraspecific variation under contrasting environmental conditions16,64. Indeed, Fitzky et al59. highlight that restricting the contribution of urban GI to O3 formation is further complicated by uncertainty in the influences that new plants will bear on future urban environments. It is projected, for example, that plants selected for contemporary urban environments will reach maturity when anthropogenic NOx concentrations are greatly reduced by the prevalence of cleaner transport, rendering bVOCs defunct as a precursor of O3 or, conversely, increasing the sensitivity of urban ozone chemistry to bVOC emissions59. Advanced air chemistry transport models that accommodate physiological models will be useful in describing bVOC-NOx-O3 relationships at different spatial scales, but such approaches to date have been incomplete59.
Pragmatic methods and principles, which encourage an overall restriction of bVOC emissions without absolute exclusion of certain plants, are therefore required (e.g. Simpson and McPherson67). Given that the majority of species are low bVOC emitting, with many presenting negligible or even indiscernible emissions64, the omission of species known to be generally high bVOC emitting may be sufficient for appropriate urban GI selection in the United Kingdom. However, this consideration is primarily for large-scale (i.e. city- or regional-scale) planting initiatives28,65. At local scale, plant diversity with respect to existing or planned GI in surrounding local areas is considered to be more important for long-term resilience under a changing climate and urban landscape7,68.
Pollen
Pollen collectively describes individual pollen grains, which carry the male reproductive cells of a plant. In angiosperms, pollen is produced in the anther, whereas gymnosperms produce pollen in the male cone. Human allergic reactions to airborne pollen are 20% more likely in urban areas than in rural areas, due in part to species homogeneity and interactions between pollen emissions and existing air pollutants69,70. Cuinica et al.71 exposed pollen from three tree species (Betula pendula, Carpinus betulus and Ostrya carpinifolia) in vitro to two low levels of NO2 (~0.034 and ~0.067 ppm), and found an associated increase in human allergenicity, as well as a significant decrease in pollen fertility. In a similar study, concerning the same species, Cuinica et al.72 found that exposure to low levels of CO, O3 and SO2 can also increase the allergenicity of pollen, to varying degrees in respect of pollutant type, exposure time and plant species. These findings supplement numerous studies that indicate that the viability and chemical composition of pollen are altered by exposure even at very low levels73. However, it also appears that pollen is less susceptible when exposed to pollution in vivo, rather than in vitro, and that the susceptibility of pollen differs among plant species73. It may therefore be inferred that judicious plant selection can, to some degree, limit pollen corruption. However, further research into the influences of pollution on pollen dispersion and allergenicity is required, particularly under climate change and contrasting environmental scenarios70,73.
Male cultivars of dioecious species that rely upon wind pollination comprise the most prolific source of atmospheric pollen from woody plants41,74. The selection of species that do not fit this profile is therefore recommended for planting programmes that aim to improve air quality, particularly where vulnerable people (e.g. schoolchildren) spend their time74. This may include the selection of female cultivars or insect-pollinated species (although insects can struggle to pollinate in polluted areas75). However, it should be noted that species diversity is paramount, not only in order to support ecosystem resilience and multifunctionality but also to minimise the emissions potential of any individual species41.
A comprehensive guide to landscaping for limited allergenic pollen emissions is provided by Ogren74. This guide includes the OPALS (Ogren Plant Allergy Scale) system, which currently ranks >3000 plants on a scale of 1 (least allergenic) to 10 (most allergenic) and according to >130 positive and negative factors. Ogren74 anecdotally suggests that tall vegetation barriers on the windward side of properties may be used to limit the exposure of inhabitants to allergenic pollen. Such vegetation barriers should be constructed of species with low OPALS ratings, which excludes many species that are commonly used for hedging, including Arborvitae spp., Cupressus spp., Ligustrum spp., and male cultivars of Juniperus, Salix and Taxus spp.74.
Advantageous aspects of GI for air quality
The primary mechanisms by which vegetation may be considered to improve air quality are those concerning dispersion and deposition29. Dispersion involves the transportation and dilution of pollutants from the pollutant source, and the various roles that vegetation may play in this process were outlined earlier (see ‘Local scale’ subsection). Dry deposition describes the process by which pollutants are deposited on solid surfaces, thereby reducing ambient atmospheric concentrations. It is through its influences on deposition that vegetation may be seen as passively filtering pollutants from the ambient air. The capability of vegetation to do so is relatively high in light of its high surface area and complexity in comparison with, for example, grey infrastructure29,76. However, the potential capacity of a species for pollutant deposition is determined by the quality and sum of its individual traits36,37,77,78.
Numerous review articles and technical papers have discussed the dry deposition process, including deposition schemes used in modelling systems, as well as methods used to describe or define deposition, deposition velocity and deposited amount29,48,79,80. Significant points regarding the influences of GI on dry deposition are summarised in Supplementary Section S1; the key message is that the influence of plant variation on deposition is largely determined by related variation in plant macromorphology and in the range of traits (including leaf physiology and micromorphology) that contribute toward surface resistance (Supplementary Section S1).
The immediate physical environment also influences the value and optimal form of GI as a pollutant sink. Under field conditions, dispersion bears an inextricable influence on deposition20,77,81. Recognising this, a number of recent studies have employed various integrated dispersion–deposition approaches to investigate impacts of GI52,82,83,84,85. For example, Morakinyo and Lam86 used such an approach in a CFD-based assessment of interactions between PM concentrations and vegetation barriers. The authors concluded that there may be trade-offs between vegetation barrier design for enhanced deposition and for enhanced dispersion86. Given the comparatively modest effects of deposition82,84,87, dispersion should be prioritised under such circumstances. However, where GI is appropriately positioned and managed, deposition is an important additional mechanism87.
Vegetation traits for enhanced pollutant deposition
Because the type and size of pollutant determine its means of deposition (Supplementary Section S1), different vegetation traits will be most effective for the deposition of different individual pollutants. A majority of studies have explicitly focussed on PM, and often only on particles >1 µm (Supplementary Table S1). Further research into the influences of individual plant traits on sub-micrometre PM and individual gaseous pollutant concentrations is highly recommended. However, empirical evidence from field experiments to date supports the generalisation drawn above that dry deposition to GI is influenced by specific traits, or inherited biophysical characteristics, the most significant of which concern leaf surface area and leaf properties or functions32,36,88,89,90. As mentioned, generic recommendations regarding plant selection for pollution (particularly PM) abatement are provided by previous works (e.g. Yang et al.37, Grote et al.77 and Sæbø et al.36). Appropriate species choice may also be gleaned from outputs generated by ecosystem service forecast models, such as the increasingly popular i-Tree tools. However, such models and associated outputs currently offer limited applicability at local scale48.
Yang et al.37 developed a ranking system for 100 tree species and evaluated each according to presumed PM2.5 removal efficiency, potentially detrimental aspects, and environmental tolerances. Although the methodology used by Yang et al.37 has limitations (see ‘Coordinated recommendations for enhanced vegetation barrier design’), the included set of determinant traits (particularly leaf complexity, size, surface feature and plant type (evergreen/deciduous)) is generally supported by findings from other studies. For example, Weerakkody et al.91 found a number of beneficial leaf traits for PM capture, including, most significantly, small size, complex shape and hairy or waxy surface. Earlier, Grote et al.77 found that foliage longevity is among the most important traits for air quality, along with canopy density, water-use strategy (an element of environmental tolerance) and bVOC emissions.
Foliage longevity and leaf phenology
Foliage longevity describes the length of time that a plant remains in leaf. Evergreen species retain functional leaves throughout the year, whereas deciduous species exist without functional leaves for part of the year and typically during the winter or dry season. The foliage longevity of deciduous species varies between and within species and is influenced not only by genotype but also by environmental conditions at microscale92. Due to the significance of leaves in pollutant deposition, the length of time during which GI may be most influential upon deposition is determined by its foliage longevity. In terms of deposition, evergreen species are therefore preferable to deciduous species, and deciduous species that generally exhibit longer in-leaf seasons are preferable to deciduous species that generally exhibit brief in-leaf seasons20,36,93. However, evergreen species may be more susceptible to certain stressors (e.g. climate warming94) than deciduous species, with potential implications for sustainable ecosystem service provision.
Leaf phenology describes the timing of leaf emergence and senescence and is also subject to inter- and intraspecific variation according to genotype and environmental conditions92. Indeed, variation is such that some species (e.g. Faidherbia albida) exhibit ‘reverse’ leaf phenology, where emergence occurs when senescence would typically occur95. Such a feature may be useful in mitigating air pollution during wintertime, when urban pollutant concentrations are often highest36,42, without restricting the palette of potential species to those that are evergreen. Given a comprehensive understanding of local-scale pollutant fluxes over time, it may even be possible to select species according to the most suitable leaf phenology. However, the selection of a species according to such narrow criteria would be to disregard the complex relationship between GI and air quality, as well as complementary services provided by GI, such as microclimate regulation during warmer seasons9. Furthermore, due consideration should be given to the suitability of each species to the anticipated environmental conditions. Many Pinus species, for example, are effective for PM deposition during wintertime, but susceptible to damage by salt spray from winter road conditioning, limiting their viability for many roadside locations36. The employment of alternative evergreen species that have exhibited tolerance to salt spray (such as Taxus spp.) may therefore be more appropriate at immediate roadsides36.
Density and porosity
The influence of a vegetation barrier on the movement of polluted air is determined by its density or, inversely, its porosity34. Abhijith et al.20 and Janhäll29 both summarise research findings on the effects of leaf area index (LAI), leaf area density (LAD) and vegetation porosity on pollutant concentrations. LAI is a dimensionless metric that describes the total leaf surface area per unit ground area (m2/m2)20,29. By contrast, LAD describes the total one-sided leaf area per unit canopy volume (m2/m3)20,96. LAI and LAD are the primary density parameters used to describe the available vegetation area for deposition29, and one or the other parameter is often used to estimate barrier porosity34. Despite the uncertainty introduced by this estimation method, and the lack of comparability between results from different studies due to variation in methods used29, a number of generalisations may be drawn regarding the influences of porosity on downwind pollutant concentrations.
Where vegetation is healthy its density is determined by leaf and branching morphology41, which varies greatly between species. A recent field investigation into the influence of GI in open-road conditions on PM10, PM2.5, PM1, BC and particle number concentration (PNC) found that downwind pollutant concentrations generally reduced with an increase in LAD32. This inverse correlation is corroborated by results from a CFD modelling study undertaken by Tong et al.55, which compared the influences of six vegetation and/or solid barrier configurations on near-road particle pollutant concentrations. Tong et al.55 found that wide vegetation barriers with a high LAD, as well as vegetation–solid barrier combinations, significantly reduced downwind particle concentrations. However, results from these studies32,55 suggest variation in concentration reduction efficiencies between particle sizes, which substantiates the conclusion by Steffens et al.97 that although reductions generally increase with the LAD of a barrier, the responses are non-linear. Similarly, based upon results from various studies, Abhijith et al.20 found that an optimal shelterbelt porosity may lie within the range of 20–40% for TSP and 10–20% for PM10, implying that optimal porosity is partly determined by target pollutant.
A higher vegetation barrier density generally results in lower downwind concentrations of both PM86,98,99 and gaseous pollutants54,100. However, CFD modelling undertaken by Ghasemian et al.54 found that a LAD of 3.33 m2/m3 reduced downwind concentrations of roadway emissions (represented by a tracer gas) by 10%, whereas a LAD of 1 m2/m3 increased downwind concentrations by 15%. These findings highlight that due to fluent penetration by pollutants and the introduction of the windbreak effect, very low-density (or very porous) vegetation barriers increase downwind pollutant concentrations until a critical point is reached at which further increases in LAD reduce downwind pollutant concentrations54. On the other hand, at very high densities (or low porosities), vegetation barriers can act as an impermeable obstacle, directing air above and around the barrier and restricting opportunities for pollutant deposition29,34. Therefore, optimal vegetation barrier density for deposition may be seen as permitting infiltration without throughflow101.
Abhijith et al.20 summarise that wide, tall and low-porosity vegetation barriers reduce downwind pollutant concentrations. The individual influences of these parameters (height, width and porosity), however, are relative not only to each other but to external factors. For example, meteorological conditions affect the porosity of different vegetation types in divergent ways; results from several studies indicate that, under high wind velocity, broadleaved and coniferous vegetation exhibit a decrease and an increase in porosity, respectively20,29. This reiterates the contextual nature of optimal porosity or density, as with all other aspects of GI. Barrier height, which should generally increase with distance from the road20, may be determined according to plant species choice or by planning requirements; barrier width is limited to available planting space; and optimal density can vary somewhat with barrier width34. In respect of the non-linear relationship between downwind pollutant concentrations and increasing LAD54,97, as well as the importance of semi-permeability101, optimal barrier density may be broadly described as above average density but below absolute. Given that the LAD of most naturally occurring vegetation is 1–5 m2/m3 102, and in line with findings by Ghasemian et al.54, a LAD of ~4 m2/m3 may be recommended. Indeed, such a value would be in agreement with recent findings from CFD modelling undertaken by Deshmukh et al.53, which suggested that a LAD of 3 m2/m3 or higher is needed for downwind PM reductions. However, it should be noted that the density of a species that forms part of a barrier under active management may differ considerably from that exhibited by the same species under ‘natural’ or comparatively unmanaged conditions. Furthermore, the porosity of two barriers that contain the same species and are managed in the same way will differ according to site conditions (light availability, microclimate, soil conditions, root space, etc.). Therefore, it may be more feasible to regulate the porosity of a vegetation barrier by post-planting management interventions (e.g. thinning a hedge that is too dense) than by species choice alone, on the condition that the species chosen has a propensity to reach the required density (i.e. naturally exhibits at least a moderately dense crown).
Leaf size and shape or complexity
Differences in leaf size and complexity are significant predictors of deposition, particularly for PM91. Numerous field experiments have found that species with smaller leaves tend to be more effective than species with larger leaves, which may be partly attributed to the higher perimeter/surface area ratio of smaller leaves103. For example, a recent study into leaf characteristics for traffic-related PM (PM1, PM2.5 and PM10) capture found that leaf size inversely correlated with PM accumulation, whereas there were no leaf surface characteristics that showed a clear correlation with PM accumulation across species104. Interestingly, this study found that the needles of Juniperus chinensis were the most effective leaves, despite being without leaf hairs or rough surfaces, followed by small-leaved broadleaf species104. This finding is supported by results from similar studies, which also found that coniferous (needleleaf) species generally offer higher deposition velocities than broadleaf species36,37,102,105,106. Sæbø et al.36 suggest that the long and narrow shape of needle leaves offer a thinner quasi-laminar boundary layer than that of broadleaves, which offers comparatively less resistance to deposition via a shortened diffusional path length107. Chen et al.105 concur that many conifers are generally more effective for PM accumulation and post-rainfall re-capture due to their acicular, needle-like shape.
For broadleaf species, Chen et al.105 found that leaf shape and venation did not affect PM deposition, but that micromorphological surface characteristics did. However, Leonard et al.78 found that leaf shape does influence PM deposition, albeit to a lesser extent than leaf surface characteristics. Indeed, a number of studies have found that complex leaf shapes are generally more effective than simple leaf shapes78,89,90,91,107. In an evaluation of leaf traits for PM deposition, Weerakkody et al.91 erected experimental rigs containing both synthetic and natural leaves alongside a busy road, including synthetic leaves of different shapes but with identical surface areas and surface characteristics. Weerakkody et al.91 found that complex (lobed) leaf shapes demonstrated a greater potential for PM deposition than simple (elliptical or linear) leaf shapes. The authors suggest that differences in the effectiveness of leaf shapes are related to the influences of leaf shape on air patterns around the leaf, as a result of the variable drag forces upon the leaf due to wind conditions91. The comparatively poor accumulation on elliptical leaves, despite their larger perimeter, is explained by their pliancy under wind flows and lower levels of associated turbulence, indicating that the rigidity of needle leaves may be significant in their effectiveness for deposition91. A comparison of results from several wind tunnel studies concerning particle capture efficiencies of different species, formulated by Conroy et al.108, found that Pinus nigra and Cupressocyparis leylandii were the first and second (respectively) most effective species under wind speeds of 3 m/s, but that Pseudotsuga menzeisii was the least effective species, behind nine broadleaf species. These findings highlight that it cannot be concluded that needle leaves are necessarily more effective for deposition than broadleaves. However, for PM deposition, it may be generalised that needle-like and small, complex leaves are more effective than other leaf types. Further research into the comparative effectiveness of different coniferous leaf shapes (needle-like, awl-shaped, scale, etc.) is highly recommended.
Leaf surface features
Several leaf surface features are considered to positively influence deposition. Plants whose leaves present one or more of these features, as well as any of the above-described biophysical traits, are therefore considered to be relatively effective. However, functional traits may vary substantially not only between but within species, according to microclimatic and environmental conditions77,92. Additionally, the functionality of any individual trait under any given circumstance is subject to internal or plant-specific factors, such as phenology, as well as external factors, such as ambient temperature109.
Generally, rough leaf surfaces are more effective for deposition than smooth surfaces36,88,91,103,107,110,111. For example, Weerakkody et al.91 reported a number of beneficial traits for PM capture, including, most significantly, small leaf size and complex leaf shape (macromorphological features, discussed above), as well as trichomes, epicuticular wax and surface ridges (micromorphological features). An earlier study undertaken by Zhang et al.111 utilised atomic force microscopy and scanning electron microscopy to examine influential leaf features for particle capture, and found the same micromorphological features to be significant for deposition. However, Zhang et al.111 observed a difference in significance of individual features between broadleaved and coniferous species: increased leaf micro-roughness (characterised by grooves and ridges on the leaf surface) was found to correlate with increased deposition to broadleaves, whereas stomatal density and amount of epicuticular wax were found to positively correlate with deposition to coniferous (needle) leaves. These findings were corroborated by a similar, subsequent study112.
Epicuticular wax ultrastructures were also found to be significant in PM deposition to both conifers and broadleaves by a number of other studies36,90,103,113. Grote et al.77 suggest that this may be partly due to leaf wettability, with particle concentrations significantly reduced through reactions with wet surfaces, and water-soluble pollutants such as NO2 and SO2 subject to dissolution in a water film at the leaf surface. However, the influence of wax structure and chemical composition on variation in deposition is not well understood and requires further research113,114.
There is also uncertainty in the comparative significance of stomatal traits, with disagreement between findings that indicate the importance of stomatal density (e.g. Zhang et al.111) and those that indicate the importance of stomatal size (e.g. Liang et al.115). Stomatal uptake particularly concerns the diffusion of gaseous pollutants from the air as a result of plant photosynthetic and water management processes116, although particles <2 µm can enter the stomatal cavity110. Stomata open and close in response to environmental conditions, and air pollutant removal may be enhanced by the selection of species with extensive opening periods (anisohydric species, which include Populus and some Quercus species) rather than species that shorten their stomatal opening period in response to drought (isohydric species, which include Pinus and Platanus species)77. However, the benefit of plants that exhibit the appropriate water-use strategy may be offset in some cases by increased susceptibility to harsh urban conditions (see ‘Environmental tolerance’).
Regarding the above results by Weerakkody et al.91 and Zhang et al.111, a link between leaf grooves and/or trichomes and effective particle capture is well-supported by the literature36,88,90,105,113,117,118. Chen et al.105, for example, found a clear correlation between PM2.5 accumulation and grooves and trichomes in leaves. This is supported by a chamber study, which compared deposition velocities across five plant species (see Supplementary Table S1) and found that certain leaf features, including grooves and trichomes, improved particle capture for (NH4)2SO4117.
Both Weerakkody et al.91 and Leonard et al.78 conclude that trichomes are the most effective micromorphological feature for particle deposition. Liang et al.115 found instead that groove proportion and stomata size positively correlate with PM2.5 capture amount, whereas trichomes and stomatal density were not found to be significant. Disagreement between these and other findings highlights that the single most influential surface feature across leaf types (broadleaf, needle-like, scale-like, etc.) for individual pollutant types is currently unclear and an area for further research118. However, it may be generalised that rough leaf surfaces (including grooves, ridges and trichomes) are more beneficial than smooth surfaces for particle deposition, that epicuticular wax is beneficial (particularly for deposition to conifers111,112) and that stomatal traits play a significant role that warrants further investigation. These points conform to different processes outlined in Supplementary Section S1, which highlights that different surface features will vary in influence for different target pollutants.
Findings from a recent study undertaken by Zhang et al.119 indicate that genetic modification may enable the selection and enhancement of micromorphological features for air pollution removal, as previously suggested by Lawson and Blatt116. Due to their complex leaf surface micromorphology, broadleaf species are more effective for deposition than coniferous species per leaf area, whereas coniferous species are more effective at tree scale due to a larger total leaf area110,115. However, Tiwary et al.120 illustrate that deposition does not necessarily increase with leaf area for all pollutant types, that a range of different processes, counter-processes and feedback mechanisms are at all times in operation, and that holistic species assessment systems are therefore required.
Source: Ecology - nature.com
