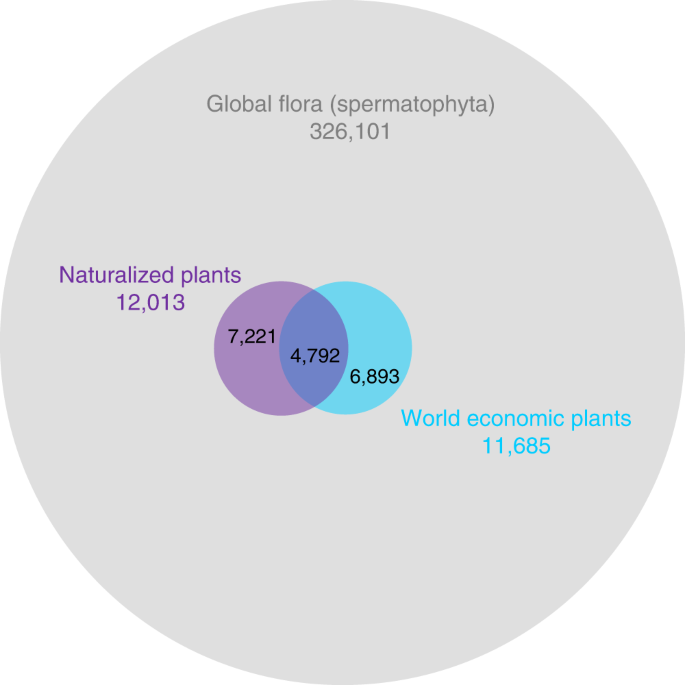
By combining the largest currently available databases on economic uses and naturalization success, we show how the varied uses of plants by humans contribute to their ability to establish outside of their native ranges. The percentage of naturalized taxa among economic plants is almost 18 times higher than among the global seed plant flora without economic uses (41.0 vs. 2.3%, Fig. 1), indicating that intentionally introduced alien plants are highly likely to become naturalized. Furthermore, we show that the likelihood of naturalization increases with the number of economic uses. Our analyses further show that naturalization success varies among economic uses, with taxa grown as animal food or for environmental uses, contributing disproportionately more to the global naturalized flora. For species with multiple economic uses, the specific combination of uses also mattered, and particularly if a focal economic use was combined with an environmental one, the likelihood of naturalization was always high. While taxa originating from the Northern Hemisphere are generally overrepresented in the global pool of economically used plants, those from temperate and tropical Asia are most likely to be naturalized in other continents. The percentage of economic plants in regional naturalized floras declines with latitude. Finally, we showed that the phylogenetic structure in naturalization success is partly, but not fully, caused by a phylogenetic bias in the plants that have been selected by humans for economic use. Altogether, our results point to a strong role of economic use of plants in driving global plant naturalization patterns.
The large proportion of economic plants that have become naturalized strongly suggests that cultivation for economic use is the major pathway for the introduction of naturalized alien plants in regions across the globe. In the global naturalized flora, 39.9% of the taxa are known to have an economic use. Strikingly, however, this percentage is substantially higher in the vast majority (98.5%) of the regional naturalized floras, with a median of 70.2%. This deviation indicates that many regions around the world share the same naturalized plants with economic uses, and thus that they differ more in the naturalized plants without economic uses. In line with this, the median number of regions in which economic plants have become naturalized exceeds the median number of regions in which plants without economic uses have become naturalized (8 vs. 3 regions; Supplementary Fig. 2). An explanation could be that most of the naturalized plants without economic uses have been introduced accidentally (e.g., as contaminants of imported grain, wool, or fibers), a process that is likely more stochastic and spatially sporadic than the directed and repeated introduction of economic plants. Overall, these findings indicate that economic plants disproportionately contribute to global biotic homogenization.
All 12 categories of economic uses have proportions of naturalized taxa that by far exceed the percentage of the global flora that has naturalized (3.7%). For bee plants and plants used for the production of non-vertebrate poisons, over 70% of the taxa have become naturalized, and for another eight categories, the percentage is over 50%. The proportion of naturalized taxa is the lowest for gene sources used for genetic improvement of crops (27.3%), most likely because these taxa are not cultivated in large numbers. The high proportion of naturalized taxa in some of the economic use categories might be explained by most of their taxa having additional economic uses. For bee plants and non-vertebrate poisons, more than 90% of the taxa have an additional economic use, whereas for gene sources this is the case in less than 30% of the taxa. So, it could be that some economic use categories have high percentages of naturalized plants because their taxa frequently have multiple uses. When we focused on the taxa that have only a single economic use—to get an unbiased estimate of their contributions to naturalization success—then the categories animal food and environmental use had the highest proportions of naturalized taxa. Animal food plants are used as fodder or forage (Supplementary Fig. 11), which are usually grown in large quantities, and repeatedly, to feed the large global populations of livestock47. Environmental plants include predominantly taxa used as ornamentals (Supplementary Fig. 11), which are likely to be grown in many gardens around the world. The other, smaller subcategories of environmental uses, such as agroforestry, erosion control, and soil improvers, have even larger proportions of naturalized taxa than the ornamental subcategory (Supplementary Fig. 11), most likely because those plants are frequently directly planted into seminatural landscapes. So, plants in the animal food and environmental categories have contributed particularly many taxa to the naturalized floras, most likely because they are widely and repeatedly cultivated in large numbers, and thus benefit from a high propagule pressure.
Both the likelihood of naturalization and the number of regions, in which an economic plant is naturalized, increased with the number of economic uses. A plausible explanation for this is that planting frequency, and thus propagule pressure, is higher when a plant has multiple uses. A similar result was reported in a study analyzing the factors that determine the naturalization of Central European species in North America—species that were utilized for a wider range of purposes by humans were more likely to naturalize27. This effect was manifested via the effect on residence time as species with a broader range of economic uses were likely to be introduced earlier in history, and thus had more opportunities to naturalize27,48. It has also been shown that species with multiple introduction pathways are more likely to have negative impacts49. So, the more uses a plant has, the more likely it is that it will naturalize.
In addition, we found that it also matters which combination of multiple economic uses a plant has. Among the taxa with two economic uses, the likelihood of naturalization was in most cases largely increased when an economic use is combined with an environmental one, and decreased when combined with the use as material. This provides further support for our conclusion that environmental plants and material plants have high and low naturalization probabilities, respectively. Most plant taxa in the subcategories of the category materials, such as the largest subcategory wax, are not cultivated very widely, and were possibly selected for specific qualities rather than for a high yield. The only exceptions were plants in the subcategories fiber and wood, which are planted more widely, and have relatively high likelihoods of naturalization (Supplementary Fig. 11). Having two (or more) economic uses resulted in a higher probability of naturalization overall, and the magnitude was mainly determined by additive effects of the economic uses. However, there were also diminishing returns of having more than one economic use, and the magnitude and significance of these interactions depended on the specific combination of economic uses (Fig. 4). So, although the likelihood of naturalization increases with the number of economic uses, the specific combination of the uses matters.
Relative to the size of the native floras, taxa from the Northern Hemisphere (temperate Asia, Europe, and North America) and Africa were overrepresented among the economic plants. Given the overall high likelihood of naturalization of economic plants, this pattern of disproportional overrepresentation could explain why the Northern Hemisphere continents are overrepresented as donors of naturalized alien plants generally39. In other words, Northern Hemisphere plants might not have a higher innate ability to naturalize, but are simply more likely to have been introduced elsewhere for economic use. If one only considers the economic plants that continents have donated, temperate Asia, as well as tropical Asia, is still overrepresented as a donor of naturalized plants in most continents, suggesting that many Asian plants have a high innate naturalization potential. However, Africa, Europe, and North America are then actually underrepresented instead of overrepresented in the naturalized floras of most other continents. This strongly suggests that the overrepresentation of European and North American plants in naturalized floras globally39 is more likely due to their higher likelihood of being introduced elsewhere in the world (as economic plants) than due to a higher innate naturalization potential of plants from those continents. The reasons for differences in innate naturalization potential of taxa from different continents and how this potential might change in the future, due to, e.g., climate change, require more research.
The proportion of economic plants in regional naturalized floras is generally higher than the proportion in the global naturalized flora, but also varies among regions. Island floras usually have larger proportions of naturalized and invasive aliens than mainland regions37,38. While this pattern may reflect a reduced resistance against introduced aliens within ecologically simple and competitively naïve island communities50,51, we had also expected that it could partly be driven by the need of humans to import alien plants into relatively species-poor island systems in order to meet economic needs. However, the proportion of economic plants in naturalized island floras was not higher than in naturalized mainland floras. Apparently, in addition to the intentionally introduced economic plants, most islands also receive many unintentional introductions of alien species. We furthermore showed that the proportion of economic plants in the naturalized floras increased toward the equator. In other words, tropical regions usually have higher proportions of economic plants in their naturalized floras than temperate regions. A potential explanation could be that the species-rich tropical regions, which have stronger biotic interactions52, have a higher resistance against invasion by alien organisms53, and that a constantly high propagule pressure and specific traits allowed some of the economic plants to overcome such strong resistance. Alternatively, it could be that unintentionally transported species (i.e., noneconomic plants) are less likely to be introduced to many tropical regions due to a less strong integration into global trade networks44.
Among the extant global seed plant flora, we found a strong phylogenetic structure in naturalization success. This is in line with the results of a previous study showing that certain plant families (e.g., Poaceae) are strongly overrepresented in the GloNAF, and other families (e.g., Orchidaceae) are strongly underrepresented37. This suggests that certain taxa in certain clades have higher naturalization potential than others. However, the phylogenetic bias in naturalization success may partly be caused by phylogenetic bias in the introduction of alien taxa. Indeed, among the extant global seed plant flora, we also found a strong phylogenetic structure for economic use of plants, and among the economic plants for most of the individual economic use categories. This indicates that closely related taxa are likely to share characteristics that make them suitable for economic use, or that they have similar histories of being brought into cultivation. When we corrected our test of phylogenetic structure in naturalization success for the economic use of plants, we indeed found that the phylogenetic structure, although it remained highly significant, became weaker. This shows that at least part of the phylogenetic patterns in naturalization success can be explained by phylogenetic biases with regard to the plants that humans use economically.
In conclusion, our global-scale analysis provides strong evidence that economic use in general, as well as the number and nature of economic uses, has strong effects on naturalization success. In particular, plants grown for animal food or environmental purposes (the latter represented mainly by ornamentals), which are most likely to be cultivated widely and in large numbers, have the highest likelihood of naturalization. Changes in the extent of cultivation may thus also affect future naturalization patterns. For example, the increasing cultivation of biofuel crops54 may increase the naturalization success of plants in this economic use category.
While it has been suggested that European plants are successful invaders in many other parts of the world due to their innate invasiveness42, our results suggest that their success is largely, though not exclusively, driven by their overrepresentation among economic plants. On the other hand, plants from temperate Asia are disproportionally overrepresented among economic plants, and also appear to have an innate greater naturalization potential. Furthermore, although the high naturalization success of some phylogenetic clades is likely to reflect innate naturalization properties of the species, our results indicate that phylogenetic biases in the economic use of plants partly underlie the phylogenetic pattern in naturalization success. Taken together, our results suggest that to fully unravel the drivers of naturalization success of plants, we must take into account their unique relationships with humans, and specifically how they are used in global economic activities. With ongoing climate change, it is likely that in many regions there will be a turn over in the economic plants that can be grown outdoors, as well as in the demands for certain plants (e.g., for biofuels). The resulting changes in the cultivation of economic plants are likely to determine future naturalization patterns.
Source: Ecology - nature.com
