Study area and design
We conducted this study within the geographical distribution centre of Swiss stone pine in the Central Alps. We chose two elevational gradients close to Davos, Switzerland, encompassing the whole elevational distribution of Swiss stone pine; one in the Sertig valley (46°44′0.76″N, 9°51′3.5″E) and one in the Flüela valley (46°48′0.25″N, 09°54′15.38″E). The forest structure at the lowest elevational belts (about 1,850 m a.s.l.) is a mixed coniferous forest, mainly composed of European larch (Larix decidua) and Norway spruce (Picea abies). The abundance of Swiss stone pine is distributed unimodally from 1,850 m a.s.l. up to 2,150 m a.s.l., where pine trees (> 3 m tall,33) form the upper tree line. Young pine trees can be found up to 2,200 m a.s.l., but none are growing at and beyond 2,250 m a.s.l.18. In each valley, we established nine elevational belts spaced by 50 m of altitude ranging from 1,850 to 2,250 m a.s.l. reaching across and beyond the elevational distribution of the pine.
Demographic processes
Seed deposition sites (i.e., seed caches deployed by Spotted nutcrackers) were recorded by randomly selecting a 2 × 10 m plot within each elevation belt. Each 20 m2 plot was composed of 20 1 m2 subplots, resulting in 20 subplots at each of the nine elevational belts per valley. In the centre of each subplot, we took soil samples (1 dm3 per sample) and thoroughly searched for deposited seeds. Intact seeds or seed shells handled by nutcrackers were recorded as cache presence and marked as a seed deposition site. We recorded seed deposition during the main seed-caching season in mid-August until beginning of September over six years (2012–2017), resulting in a total number of 2,156 soil samples. In these soil samples, we found 256 (12%) seed caches deposited by nutcrackers across the elevational gradient in both valleys and all years.
To determine rates of seed predation, seed germination and seedling survival, we conducted a seed transplant experiment across the nine elevational belts. According to a random-stratified sampling design, we selected five microhabitat types at each elevational belt for the experiment (1. soil covered by vegetation, 2. close to adult Swiss stone pine individuals [i.e., up to a distance of 1 m], 3. open soil, 4. rocky habitat, 5. microsite covered by snow [i.e., late snow lie areas]). For elevational belts above the tree line (at 2,250 m a.s.l.), the microhabitat “close to adult Swiss stone pine” was replaced by matgrass (Nardus stricta L.) dominated sites, to guarantee an equal sample size in each elevational belt. At the beginning of the growing season (i.e., end of May), we placed Swiss stone pine seeds in a plastic mesh at two to six replicates per microhabitat at each elevational belt, resulting in a total number of 1,980 seed sowing replicates, including 6,858 seeds monitored during the study (for detailed information on the number of replicates deployed per year see Supplementary Table 3). Each mesh (i.e., insect gauze with 1.5 mm holes, open at the top) contained five Swiss stone pine seeds simulating the average number of seeds per cache deposited by nutcrackers16. From the total number of 1,980 meshes, 540 meshes were protected by 1.5 mm wire-cages in the field to prevent loss of seeds. Meshes were buried about 4 cm deep in the soil and fixed with metal pins. To break dormancy of the seeds, seasonal variation was simulated in a wet clay-sand mixture by exposure to temperature shifts between 5–25 °C for 22 weeks. At the end of the growing season (i.e., end of September), we evaluated whether seeds had been preyed upon or germinated. Further, we monitored the survival of seedlings until the end of the subsequent growing season in the following year. Out of 6,858 seeds monitored in the six study years 3,023 seeds (44%) were preyed upon and removed by rodents and other animals and 451 (7%) germinated within the first growing season (i.e., the period between May and September). Of 319 seedlings 65 (20%) survived to the end of the following growing season (i.e., the period from September to September of the following year) in two years (i.e., 2014–2015, 2015–2016). We included only these two years of survival as these were the only years where numerous seedlings survived to the next year.
Environmental variables
Canopy and ground vegetation cover are decisive for seed caching behaviour of Spotted nutcrackers17, 18, and they strongly affect seed germination and seedling survival of Swiss stone pine via shading and abiotic soil properties (i.e., soil surface temperature and soil moisture)18 as well as the presence of fungal pathogens19. Therefore, these two factors do affect both the selection of caching sites and the subsequent demographic processes of plant recruitment of Swiss stone pine. Canopy cover was measured at each seed deposition subplot and seed sowing replicate with a spherical densitometer. On the same sites, ground vegetation cover was assessed by estimating the percentage cover of dominant ground flora species: Juniperus communis L., Loiseleuria procumbens (L.) Desv., Vaccinium spp. L. and Rhododendron ferrugineum L. within 1 m234. In total, we measured canopy cover and ground vegetation cover at 4,136 1 m2 plots, i.e., at 2,156 seed deposition subplots and 1,980 seed sowing replicates.
We characterised the frequency distribution of canopy and ground vegetation cover across three range positions. To do so, we used the data collected at the 2,156 1 m2 seed deposition subplots across the 9 elevational belts and categorized these into three range positions: (1) three lowest elevational belts (i.e., lower range margin, 1,850–1,950 m a.s.l.), (2) three central elevational belts (i.e., centre, 2,000–2,100 m a.s.l.) and (3) three high elevational belts (i.e., upper range margin, 2,150 – 2,250 m a.s.l.). The relative frequency of canopy and ground vegetation cover for each range position was determined, taking into account the interaction between the two measures. To this end, each 1 m2 subplot was assigned to a cell of a 10 × 10 grid of values for canopy cover and ground vegetation cover ranging from 0 to 100% in steps of 10%. Each combination of canopy cover and ground vegetation cover was counted as the sum of subplots within the respective combination of values.
In addition, we assessed how the microhabitat characteristics (canopy and ground vegetation cover) control microclimatic environmental conditions in terms of soil surface temperature and soil moisture. Soil surface temperature was recorded every four hours over the duration of the study using iButton data loggers (Maxim) at 1,343 seed deposition and seed transplant sites across the elevational gradient and years. We calculated the mean of daily temperatures of the hottest three months of each year (i.e., June, July, August). Soil moisture was recorded on 4,136 seed deposition and seed transplant sites across the elevational gradient and years. Soil moisture was recorded under dry weather conditions in September by averaging five tensiometer (Theta-Kit version 3) measurements at each seed deposition and seed transplant site. Using linear mixed models including canopy cover and ground vegetation cover, we found soil surface temperature during the hottest three months and soil moisture to be negatively associated with high canopy and ground vegetation cover (Supplementary Table 2). Thus, our results suggest that the microhabitat characteristics are the main cause for site by site variation in microclimatic conditions, such as variations in soil surface temperature and soil moisture. Hence, we focussed on environmental variation in canopy cover and ground vegetation cover, as the most appropriate descriptors of the variation in microhabitat conditions across the species range.
Statistical analyses
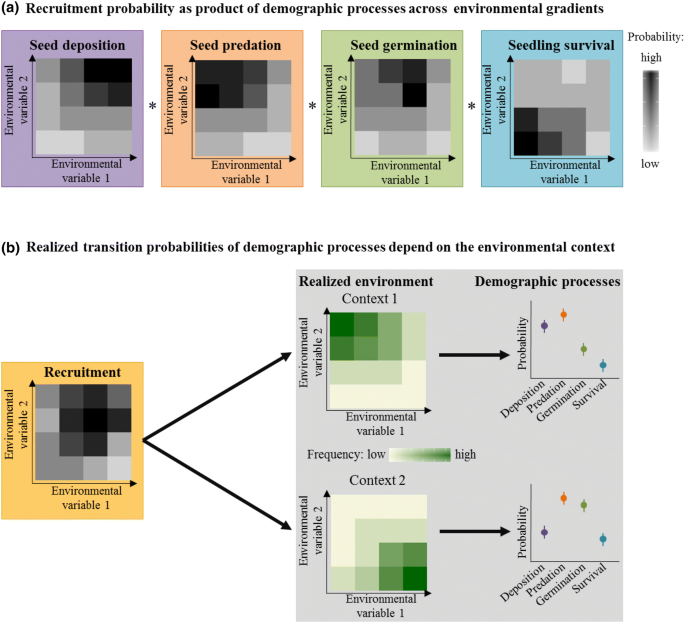
We assessed the recruitment probability across and beyond the whole elevational range of Swiss stone pine. To do so, we fitted four models describing the determinants of seed deposition (i.e., the presence or absence of cached seeds in soil samples taken at each subplot), seed predation (i.e., absent seeds and seeds that showed signs of seed predation [e.g., bite marks] at seed translocation sites), seed germination (i.e., seed germination and seedling establishment within the first growing season), and seedling survival (i.e., survival from the end of the first growing season to the end of the second growing season). We used the presence or absence of cached seeds rather than the actual seed number recorded at a caching site to account for potential methodological constraints in assessing the exact number of deposited seeds. We used generalized linear mixed models with a binomial error distribution in the R package ‘lme4′35. We included canopy cover, ground vegetation cover and their interaction as continuous explanatory variables into the models. In addition, we included plot ID, site and year as random factors to account for spatial and temporal non-independence. An observation-level random effect was included in the seed predation and seed germination models to account for overdispersion35,36. All predictors were mean centred and scaled to unit variance to allow for comparison of effect sizes across predictor variables and models. We used the models to predict the expected probability of seed deposition, predation, germination and first-year seedling survival across the gradients of canopy cover and ground vegetation cover (Fig. 2a–d). Then, we calculated the overall recruitment probability as the product of the expected transition probabilities (P) from the four individual models of each demographic process: (P_{{{text{Recruitment}}}} = P_{{{text{Deposition}}}} times left( {1 – P_{{{text{Predation}}}} } right) times P_{{{text{Germination}}}} times P_{{{text{Survival}}}}) (Fig. 2e).
We conducted a context-specific analysis to evaluate how the realized transition probabilities of the four demographic processes change at the three different range positions (i.e., lower range margin, centre, upper range margin). To account for the environmental context, we used the realized frequency distribution of canopy and ground vegetation cover at each range position to calculate the weighted mean of the realized transition probability of each demographic process and overall recruitment in the respective environment (Fig. 1). To gain an estimate of uncertainty (confidence intervals) for the realized transition probabilities, we performed a bootstrap analysis with 1,000 replicates. Bootstrap replicates were used to test for significant pairwise differences in realized transition probabilities between range positions (based on two-tailed P-values). Bonferroni correction was used to adjust P-values for multiple comparisons.
Source: Ecology - nature.com

