Fe-speciation data show pronounced water column redox stratification during deposition of the Kuibis Subgroup5. Marine transgression during deposition of the Mara, Kliphoek and lower Omkyk members resulted in basin-wide shoaling of the redoxcline, represented by dominantly anoxic ferruginous deeper and shallower (above fair weather wave base) waters (Fig. 2b,c)5,6. The data then show more frequent development of oxygenated conditions through the upper Kuibis Subgroup and the lowermost Schwarzrand Subgroup in both shallower and deeper water. However, short-lived anoxic intervals were a common feature of mid-ramp environments. After this, the inner ramp succession was dominantly deposited under oxic water column conditions, while mid and outer ramp sections were also oxygenated, at least by deposition of the upper Schwarzrand Subgroup. Furthermore, the combination of low and stable FeHR/FeT and increasing Mn/Fe (Figure S11) suggest a progressive increase in oxygenation during deposition of the Urusis Formation, culminating in conditions conducive to widespread Mn oxidation. Fossil evidence for habitation by trace making organisms occurs in all stable oxic settings from the Nudaus Formation onwards (see Supplementary Information).
Phosphorus systematics provide insight into nutrient cycling as the redox state of the water column developed in the Nama Group. In particular, consideration of P speciation and P/Al ratios helps to constrain potential productivity feedbacks arising from changes in redox conditions41,42,43. Upwelling of ferruginous deep waters may promote P drawdown in association with iron minerals, thus potentially resulting in elevated P/Al and a negative productivity feedback44. By contrast, P drawdown in association with iron minerals would be expected to be less significant under oxic water column conditions. However, whilst some ferruginous samples do show P enrichment relative to average shale (Fig. 3b), the majority are depleted, and there is no systematic difference in relation to oxic samples.
Low average P/Al in ferruginous samples of the Kuibis Subgroup may be a consequence of either generally low phosphate availability in the water column, or P recycling from sediments back to the water column. The cycling of P in sediments is largely contingent upon the redox state of the porewaters and water column, with sulfidic conditions being particularly conducive to P recycling41. Anaerobic organic matter remineralisation results in the preferential release of P, giving high Corg/Porg ratios relative to the Redfield ratio45, while reductive dissolution of Fe (oxyhydr)oxide minerals also releases P to solution46. The P released by these processes may undergo ‘sink switching’ to authigenic phases such as carbonate fluorapatite43 or vivianite47, or may be re-adsorbed to Fe (oxyhydr)oxide minerals where they persist48. However, dissolved P may also be recycled back to the water column, particularly under anoxic conditions, potentially promoting a positive productivity feedback41.
Our redox data suggest limited sulfide production during diagenesis under both oxic and ferruginous conditions (giving very low Fepy/FeHR ratios; see Supplementary Information). Such conditions would be expected to limit P recycling from the sediment, particularly under oxic conditions (where anaerobic organic matter degradation and the reductive dissolution of Fe (oxyhydr)oxide minerals are restricted), and this can be tested by considering Corg/Porg and Corg/Preac ratios relative to the Redfield ratio47. In oxic samples of the Nama Group, Corg/Porg ratios (Fig. 3e) cluster around the canonical Redfield ratio (106:1). This suggests little preferential release of P from organic matter, which would be consistent with limited anaerobic organic matter remineralization under these low TOC conditions (Fig. 3a). Relatively low Corg/Preac ratios, coupled with a high proportion of authigenic P (Fig. 3c–e), suggests that the P released from oxic organic matter degradation and the reductive dissolution of Fe (oxyhydr)oxides deeper in the sediment was subsequently fixed in the sediment via ‘sink-switching’, with no evidence for recycling back to the water column. Nevertheless, Corg/Porg ratios at the Redfield ratio argue against chronically nutrient limited productivity (which may raise primary Corg/Porg values to as high as ~60049), and instead our data suggest that the oxic Nama basin experienced rates of productivity typical of oxic marine settings.
Some ferruginous samples have elevated (above the Redfield ratio) Corg/Porg ratios, suggesting more effective anaerobic organic matter remineralization relative to oxic samples, as would be expected beneath an anoxic water column. Corg/Preac ratios are at or below the Redfield ratio, which suggests significant fixation of P in the sediment following draw down with Fe (oxyhydr)oxide minerals. However, despite this enhanced draw down mechanism, the majority of anoxic samples have significantly higher Corg/Preac ratios relative to oxic samples, which implies a degree of recycling of P back to the water column under ferruginous conditions, with the potential to stimulate a relative increase in productivity41.
A specific complication in P speciation analyses of ancient sedimentary rocks involves the post-depositional recrystallisation of authigenic apatite during burial diagenesis50. The modified SEDEX protocol employs a sequential chemical extraction which targets Pauth (using 1 M Na acetate buffered to pH 4.0) prior to extraction of Pdet (using 1 M HCl) and as such, any decrease in the solubility of primary Pauth as a consequence of burial diagenesis, will be represented by potential transfer of Pauth to the Pdet pool38. It has been noted that the detrital P content of modern continental margin sediments is 186 ± 21 ppm43, whereas modern oligotrophic settings are characterised by Pdet in the range 62–310 ppm51. Samples from the Nama Group have Pdet concentrations in the range 4.2–536.4 ppm (mean 208.7 ppm), with maximum values significantly greater than average Pdet of modern shelf environments. This may suggest that a portion of extracted Pdet represents burial recrystallisation of initially authigenic P. There is no significant correlation between Preac (as a percentage of PTot) and Al (r2 = 0.077, Figure S2c), implying negligible contamination of the Pauth pool by Pdet. Consequently, concentrations of Pauth (and by extension summed Preac) likely represent minimum values, whilst those of Pdet represent maximum values (see Supplementary Information for further information).
The combined redox proxy data suggest more oxidising water column conditions in the Nama Group basins (above that required to fully oxidise Fe(II)) in inner to mid ramp environments by the time of deposition of the lower Schwarzrand Subgroup, commencing at ~547 Ma (Fig. 2)14. The data also support a further progressive increase in oxygenation of the inner ramp to levels sufficient for Mn oxidation by the upper Urusis Formation at ~542–540 Ma13,15,52. Whilst outer ramp sections are sparse in the Nama Group, the available data show that ventilation of the outer ramp occurred, at the latest, by deposition of the Feldschuhhorn Member at ~542 Ma13,52.
What then, caused the progressive ventilation of the Nama basins at ~547 Ma? Fig. 2f shows available data of the chemical index of alteration through the Nama succession. The extent of chemical weathering is dependent upon factors including tectonics and regional climate, and affects the regional supply of nutrients from the continent and the maturity of terrigenous clays37. High average CIA values are a dominant feature of the Kuibis Subgroup, and argue for a high degree of chemical weathering, potentially mediated by hot and humid regional climatic conditions37. This is followed by a shift, in the Zaris sub-basin, to lower average values that may record a corresponding transition to cooler and drier conditions. There is a distinct peak in CIA that relates to shales of the Vingerbreek Member from the Witputs sub-basin which were deposited atop a sub-basin scale unconformity attributed to a putative short-lived regional glaciation53. This peak reflects sediment transport from a source area to the east on the Kalahari Craton and is coeval with a short-lived return to local water column redox stratification and ironstone deposition (see Supplementary Information, section 3d). The overall decrease in the CIA up-section, coupled with the observed covariation with changes in dominant water column redox conditions, point towards a driving role for changes in continentally derived nutrient influx to the basin through time. In this scenario, initial transgression and a high degree of chemical weathering led to deposition of sediments and delivery of nutrients from the Kalahari Craton to the east of the Nama sub-basins. At this time, nutrient input may have been supplemented by a degree of upwelling from the openly connected Brazilides ocean to the west, leading to elevated regional productivity and anoxia (Figs. 4a, S13). However, gradual closure of the Brazilides54 ocean likely stifled upwelling in this region, forcing the system to primarily depend upon nutrients supplied from continental weathering. The degree of chemical weathering and associated nutrient input decreased in the Schwarzrand Subgroup, possibly associated with regional climatic change53, which would have reduced primary production and hence the extent and maintenance of water column anoxia. The Fe and P speciation data provide insight into the dynamic, regional burial and recycling of the major limiting nutrient P under both ferruginous and oxic conditions11. The shallow redoxcline evident during deposition of the lower Kuibis Subgroup readily facilitated the anaerobic degradation of organic matter (and also dissimilatory Fe reduction) in sediments and likely resulted in a small degree of P recycling back to the water column, thus fuelling local productivity (Fig. 4a). However, ferruginous conditions also promoted the removal of reactive P in association with iron minerals, thus limiting the extent of this positive productivity feedback over a timescale of millions of years. By contrast, during deposition of the Schwarzrand Subgroup (547–540 Ma), P was more effectively buried in sediments through sink-switching to authigenic apatite after initial drawdown in association with organic matter and Fe (oxyhydr)oxide minerals (Fig. 4b). The lack of P recycling from oxic sediments of the Schwarzrand Subgoup stabilised oxia in the overlying water column. There is a notable increase in bioturbation intensity recorded in the lowermost Schwarzrand Subgroup55, however the potential secondary influence of bioirrigation and mixing-induced sedimentary P retention on local water column productivity remains uncertain.
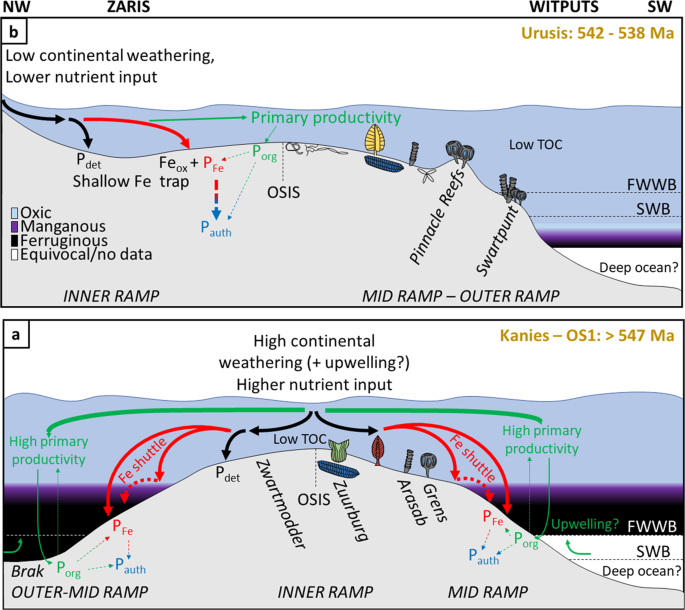
Proposed model for the co-evolution of redox, P burial mechanisms, and evolution of metazoans in the (a) lower Kuibis Subgroup (~550–547 Ma), and (b) upper Schwarzrand Subgroup (540–538 Ma). Dashed straight arrows denote either sedimentary P diffusion to the water column under ferruginous water column conditions or post-depositional ‘sink-switching’ of P.
The consistent spatial separation of anoxic deep waters from the distribution of soft-bodied, skeletal and ichnotaxa in the Nama Group may support the contention that oxygen was a metabolic requirement of these organisms. High resolution sub-sampling of fossiliferous beds in the Nama Group has previously revealed the ability of the skeletal Namacalathus to opportunistically colonise the substrate during fleeting oxic episodes under a regime of dominantly ferruginous water column conditions5. Similarly, palaeoredox studies of fossiliferous shales of the Blueflower Formation, NW Canada, have been interpreted to show the ability of soft-bodied forms including possible Pteridinium, Inkrylovia and Windermeria, to opportunistically colonise environments that were briefly ventilated for shorter durations than those resolvable by bulk Fe proxy capability56. Ferruginous conditions that dominated the deeper water column during deposition of the Kuibis Subgroup may have limited habitable space for soft-bodied macrobiota, the majority of which are preserved in the shallowest environments. However, some in situ Ernietta and Rangea specimens are also suggested to have been able to opportunistically colonise the substrate during short-lived oxic episodes, similar to the Blueflower Formation, or the ability to survive incursions of anoxic bottom waters57.
Whilst there is no paucity of siliciclastic facies in the Witputs sub-basin throughout the Nama succession, the majority of soft-bodied fossil representatives are known only from shallow, shoreface and inner-ramp, quartz-rich sandstone horizons. Particularly prominent fossiliferous quartzites appear in the Aar, Nasep and Spitskop members (see Supplementary Information Table S1 for references)23,57. However, a notable trend of increasing bioturbation intensity is present in the shallow inner to mid-ramp during deposition of the Nudaus Formation55, within the middle part of the Nama Group succession (Fig. 2g). There is a further increase in bioturbation intensity from the lower Schwarzrand Subgroup to the basal Cambrian unconformity in the Nama Group26,55. Indeed, recent quantification of bioturbation intensity in the Nama Group shows an overall increase in the percentage of bedding plane bioturbated from a mean value of 1.94% in the Kliphoek Member to 3.34% in the lower Urusis Formation, and a further increase to 5.61% in the Spitskop Member, immediately beneath the basin-wide unconformity that marks the top of the Urusis Formation (Fig. 2g)55. The increase in bioturbation intensity occurs coincident with the stabilisation of oxic conditions in this location (Fig. 2).
The local transition to stable, oxic conditions observed in the Nama succession may have been a prerequisite for the increase in bioturbation intensity in this shallow ramp setting, allowing the efficient exploitation of nutrients held within the sediment and microbial mats. However, the link between oxygenation and increased bioturbation intensity is less clear in broadly contemporaneous successions of South China. Here, sediments of the Dengying Formation contain trace fossils58 alongside proxy evidence for only punctuated oxic intervals under a regime of dominantly anoxic water column conditions9,59. Evidence from nitrogen isotope studies60 of fossiliferous Ediacaran and early Cambrian strata in South China attest to a regional nutrient regime distinct from that of the Nama Group, with high nutrient delivery to the Yangtze platform and slope, likely (in part) supplied by enhanced upwelling, driving an intense and long-lived OMZ throughout this interval9. A comparison of the relative rates of ichnotaxa diversification and their spatial distributions between stratigraphic successions of the Nama Group and South China, is required to test the relationship between palaeoredox of equivalent facies and relative diversification rates between these two potentially differing late Ediacaran palaeoenvironments.
Skeletal macrofossil assemblages of the Ediacaran attain their highest abundance and diversity in the Kuibis Subgroup of the Nama Group, where surface and mid-depths were occasionally well oxygenated but deeper waters were anoxic and nutrient rich5,6. Indeed, iron speciation data support the importance of a dominantly oxic local environment for the most diverse skeletal assemblage in the Nama basin (Driedoornvlakte, Figure S7). In the Dengying Formation, the shallowest environments of Shaanxi province and the Baimatuo Member in the Yangtze Gorges host relatively depauperate, microbial mat-related skeletal assemblages (e.g.61), where rare earth element profiles suggest continued redox instability with only short-lived oxic intervals59. The differing geochemical characteristics observed between palaeoenvironments of the Nama Group and Yangtze Platform may suggest a delicate balance between oxygen demand and nutrient loading as an incentive for the radiation of motile macrobiota, and potentially for ecosystem habitation by skeletal organisms in the late Ediacaran. This may allude to a ‘sweet spot’ in oxygen and nutrient demand for skeletonising and filter feeding ecologies in the late Ediacaran, however this remains to be tested.
While geochemical evidence from the uranium isotopic composition of carbonates (δ238Ucarb) points to an increase in the global extent of anoxic ocean waters at ~547 Ma62,63, the redox data reported here show that, on a local scale, the Nama basins became more stable and oxic at this time. These global and local datasets are not necessarily in conflict, as the uranium proxy data may record an integrated global increase in the spatial extent of productive mid-depth waters at ~547 Ma. In fact, recent compilations of δ238Ucarb from globally distributed late Ediacaran successions do suggest a trend towards an increasingly oxygenated global ocean between 545–540 Ma, in the wake of the highly negative values recorded in the lower Nama Group and Dengying Formation62,63,64. However, these global oxygenation events were transient, and their influence on the pace of animal evolution in shallow shelf ecosystems which were subject to highly heterogeneous local redox conditions demands further investigation.
Source: Ecology - nature.com

