Based on their divergent morphology (Figs. 2–5, S1–S12) and the results obtained from the molecular phylogeny (Figs. 6, S13), we describe here Jurasai digitusdei gen. et sp. nov., J. itajubense gen. et sp. nov. and Tujamita plenalatum gen. et sp. nov., for which we erect a new family Jurasaidae fam. nov. Below we provide remarks on their biology, ecology and behavior, as well as concise diagnostic descriptions for all new taxa based primarily on salient diagnostic characters. Detailed morphological diagnoses and descriptions are given in the Supplementary Text.

Live specimens of Jurasaidae fam. nov. (a) larvae of Tujamita plenalatum gen. et sp. nov. (upper specimen) and Jurasai itajubense gen. et sp. nov. (lower specimen); (b) larva of T. plenalatum; (c–e) J. itajubense: larva, female pupa and adult female, respectively; (f) adult female of T. plenalatum.

Male pupae of Jurasaidae fam. nov. (a,b) Jurasai itajubense gen. et sp. nov., dorsal and ventral view, respectively; (c) Tujamita plenalatum gen. et sp. nov., lateral view.

Mating behavior of Jurasai itajubense gen. et sp. nov. (a) male touches a female with mouthparts; (b) male touches a female with aedeagus.

Adult males of Jurasaidae fam. nov. (a,b) Jurasai itajubense gen. et sp. nov., dorsal and ventral view, respectively; (c,d) J. digitusdei gen. et sp. nov., dorsal and ventral view, respectively; (e,f) Tujamita plenalatum gen. et sp. nov., dorsal and ventral view, respectively.
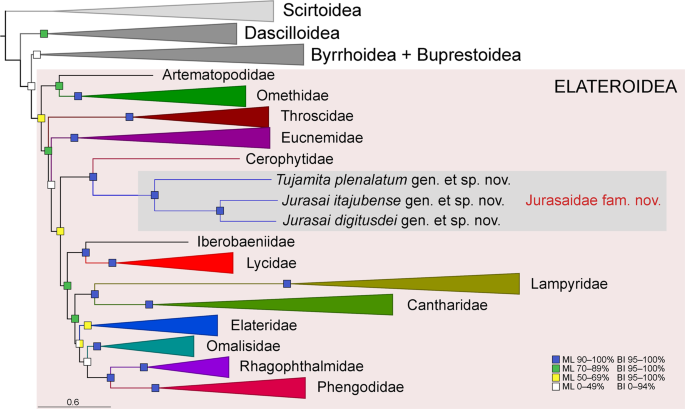
Phylogenetic position of Jurasaidae within Elateriformia based on the maximum likelihood (ML) analysis performed on the four-gene 251-taxa dataset aligned using the MAFFT algorithm. Statistical support for branches includes bootstrap values for the ML analysis and posterior probabilities for the Bayesian inference (BI). The full-resolution tree with taxon names is given in Fig. S13.
Observations of behavior of the immature stages and adults
Immature stages of two species (out of three) representing two different genera are known to date. Larvae and pupae of J. itajubense and T. plenalatum occur sympatrically in the Municipal Biological Reserve Serra dos Toledos. They are usually found in the 5–20-cm-thick layer of soil just under the leaf litter (Figs. 1c, 2a–d, 3). Although both species share the same microhabitat, T. plenalatum is much more rare; of the total 87 larvae collected, only four belonged to that species. The feeding habits of both larvae and adults are not known. Despite our offering the larvae various potential food items in the laboratory, no movement of the mouthparts or signs of pumping into the esophagus were observed. However, since many larvae were found between roots, we hypothesize that they feed on juices of fungal hyphae. Both larvae and adults usually stay in the soil. When unearthed and disturbed they become active and soon bury themselves again into the substrate. Bioluminescence is present in several elateroid lineages in Brazil, i.e., Lampyridae (fireflies), Phengodidae (glowworms) and Elateridae: Pyrophorini (luminous click-beetles)22,27,28. However, it was not detected in Jurasaidae, neither by the naked eye nor by using long exposure photography. In both species, the pupal stage lasts 22–24 days in males and 8–9 days in females. Whilst a male pupa can move only its abdomen, a female pupa is capable of walking, although it usually stays burrowed in the substrate or resting on the surface in a C-shaped position (Fig. 2d). The adult female is wingless and resembles a larva in general appearance, with only certain parts of the head adult-like (Jurasai, Fig. 2e) or with the head, prothorax and legs adult-like, though different in appearance from the corresponding parts in the male (Tujamita, Fig. 2f) (Table S1). About a week after eclosion, the male actively searches for a female and then copulates several times each for a short period of time (Fig. 4, Supplementary Video). More detailed information on field and laboratory observations are given in the Supplementary Text.
Systematics
Jurasai gen. nov
(Figs. 1d, 2a,c–e, 3a,b, 4, 5a–d, S1–S4, S6a–i, S8a,b, S10a–i, S11a,b, S12a–h).
Type species. Jurasai itajubense sp. nov.; by present designation.
Diagnostic description. Adult male (Figs. 4, 5a–d, S1, S2a–h, S4, S6a–i, S8a,b). Body length from frons to apex of abdomen 2.5–3.0 mm, from frons to apex of wing 3.5–5.0 mm. Head with antenna approximately as long as distance between head and elytral apex; labrum 1.8–4.0 times as wide as long; maxillary palpus 4- or 5-segmented; apical labial palpomere 2–5 times longer than basal palpomere. Pronotum (Figs. S2b, S4b,d, S6e) widest at anterior half, posterior half narrowed posteriad; without lateral carinae; posterior edge smooth, not marginated; mesoventrite (Figs. S4f–h, S6e) 1.1 times longer than wide, with anterior margin deeply arcuate; mesepimeron with anterior part indistinct in lateral view, separated from mesanepisternum by weakly impressed suture; mesocoxal cavities separated at middle by 1.3 times mesocoxal cavity width; metaventrite (Fig. S4f) 1.3 times longer than wide, 2.5 times longer than mesoventrite, widest at anterior 1/3. Elytra (Figs. 5a,c, S1a,f) shorter than abdomen, tapered apically, with median edges separated and divergent apicad, lateral edges sinuate, apices swollen; hind wing (Fig. S8a,b) surpassing elytral apex by 0.5–0.7 times elytral length, apical field long (0.5–0.6 times as long as total wing length). Tarsomere IV (Fig. S1e) evenly sclerotized, truncate apically. Abdomen (Fig. S1h) narrow, with sides subparallel; phallus and parameres (Figs. S2f,g, S4m–o) together 1.6 times wider than long; endophallus emerging from dorso-apical elongate notch. Adult female (Figs. 2e, 4, S2i,j, S3) (based on J. itajubense only). Abdomen and thorax almost equal to those of larvae; head with a pair of pigmented stemmata; mandible falcate; labrum free; maxilla and labium separated, labium not channeled to fit mandibles; antenna with four antennomeres; leg with tibia and tarsus separated, tarsus 1-1-1, with a pair of claws. Mature larva (Figs. 1d, 2a,c, S10a–i, S11a,b, S12a–h) (based on J. itajubense only). Body 4–7 mm long, slender, 9–10 times longer than wide; clypeolabrum (Fig. S12c) with two long setae. Mandibles and labium (Fig. S10b) forming elongate and sharpened beak-like mouthpart; mandible (Figs. S11b, S12e,f) with apex gradually sharpened apicad, parallel-sided, base elliptical; hypopharyngeal bracon hyaline. Pronotum (Fig. S12a) with four setae between lateral edge and lateral sclerotized stripe (one pair anteriorly and one pair posteriorly), two setae between stripes anteriorly; mesothorax, metathorax, and abdominal segments I–VIII (Fig. S12a,b) with one pair of laterodorsal setae near anterior margin, one pair of laterodorsal setae near posterior margin, and a pair of lateroventral setae near posterior margin. Legs (Fig. S10g) separated by 5–6 times diameter of coxa.
Etymology. From Tupi-Guarani language; Jura = mouth; saí = minuscule, thin; allusion to the
larval mouth. Gender: neuter.
Composition and distribution. Jurasai itajubense (Brazil: Minas Gerais) and J. digitusdei (Brazil: Rio de Janeiro).
Jurasai itajubense sp. nov
(Figs. 1d, 2a,c–e, 3a,b, 4, 5a,b, S1a–e, S2, S3, S6a–e, S8a, S10a–i, S11a,b, S12a–h).
Type material. Holotype, male, “Brazil, Minas Gerais state, Itajubá municipality, Biological Reserve of Serra dos Toledos (22°25′21.3″S 45°22′06.2″W), 1,358 m, soil ravine, collected as pupa on 7.VI.2018 (adult on 5.VII.2018, died on 8.VIII.2018), Rosa S.P, Barbosa T. & Paiva J. leg.” (deposited at MZUSP). For information on paratypes and other material examined see the Supplementary Text.
Diagnostic description. Adult male (Fig. 5a,b). Labrum with anterior margin emarginate; maxillary palpus 4-segmented; elytra strongly tapered; hind wing venation with only RA1+2 and MP1+2 veins; abdominal sternite VIII partly exposed; parameres with apices tapered and curved inwards. For more details see the generic diagnostic description and the Supplementary Text.
Etymology. From the type locality, Itajubá, in Minas Gerais state, Brazil.
Jurasai digitusdei sp. nov
(Figs. 5c,d, S1f–i, S4, S6f–i, S8b).
Type material. Holotype, male, “Brazil, Rio de Janeiro State: Teresópolis, Parque Nacional da Serra dos Órgãos, malaise trap, PVE 6B (22°28′11″S 43° 0′5.3″W, 868 m), VI.2015, Silveira & Khattar leg.” (deposited at DZRJ). For information on paratypes and other material examined see the Supplementary Text.
Diagnostic description. Adult male (Fig. 5c,d). Labrum with anterior margin rounded; maxillary palpus 5-segmented; elytra weakly tapered; hind wing venation with RA1+2, R3, RA3+4, RM loop and medial field veins; abdominal sternite VIII concealed; parameres with apices sausage-like, directed posteroventrad. For more details see the generic diagnostic description and the Supplementary Text.
Etymology. From Latin; digitus = finger, dei = of god; allusion to “Dedo de Deus”, a mountain peak near the type locality in Serra dos Órgãos National Park, whose shape resembles a hand pointing up towards the sky.
Tujamita gen. nov
(Figs. 2a,b,f, 3c, 5e,f, S5, S6j–l, S7, S8c, S9, S10j–m, S11c–e, S12i–l).
Type species. Tujamita plenalatum sp. nov.; by present designation.
Diagnostic description. Adult male (Figs. 5e,f, S5, S6j–l, S7, S8c). Body length from frons to apex of abdomen 3.1–4.5 mm, from frons to apex of wing 3.2–4.6 mm. Head with antenna approximately 2/3 as long as distance between head and elytral apex; labrum four times as wide as long; maxillary palpus 5-segmented; apical labial palpomere 6–8 times longer than basal palpomere. Pronotum (Figs. S5d, S6j) weakly narrowed posteriad, with lateral margins subparallel and carinate; posterior edge marginated; mesoventrite (Figs. S6k, S7f–h) 1.3 times wider than long, with anterior margin weakly arcuate; mesepimeron with anterior part distinct in lateral view, separated from mesanepisternum by grooved suture; mesocoxal cavities separated at middle by 0.7 times mesocoxal cavity width; metaventrite (Fig. S7g) 1.1 times wider than long, 2.8–2.9 times longer than mesoventrite, widest at midlength. Elytra (Figs. 5e, S5a) slightly shorter or as long as abdomen, parallel-sided, median edges contiguous to apex; apices flat. Hind wing (Fig. S8c) surpassing elytral apex by 0.2 times elytral length, apical field short (0.3–0.4 times as long as total wing length). Tarsomere IV (Fig. S5e) deeply notched. Abdomen (Fig. S5f) wide, with sides rounded and tapered apicad; phallus and parameres (Fig. S7o,p) together 1.1 times wider than long; endophallus emerging from apical oval orifice on dorsal surface of phallus. Adult female (Figs. 2f, S9). Meso-, metathorax and abdomen almost equal to those of larva; head, pronotum and leg (Fig. S9a–g) adult-like but different from those of male: compound eyes very small, flat, not protruded; posterior tentorial pits absent; antenna moniliform, with nine antennomeres; pronotal lateral carinae absent, prosternum reduced to very narrow sclerotized strip; tarsi 4-4-4. Mature larva (Figs. 2a,b, S10j–m, S11c–e, S12i–l). Body 4–6 mm long, stout, 5–6 times longer than wide; clypeolabrum (Figs S10j, S12i) with four long setae; mandibles and labium (Figs. S10j,l, S11c–e) forming short, stout, beak-like mouthpart; mandible (Figs. S11d, S12l) with apex abruptly sharpened apically, convergent anteriad, base triangular; hypopharyngeal bracon sclerotized. Pronotum with five setae between lateral edge and lateral stripe (one pair anteriorly, one pair posteriorly, and single seta at midlength), four setae between stripes (one pair anteriorly and one pair posteriorly); mesothorax, metathorax, and abdominal segments I–VIII with six setae at midlength (one pair lateral, one pair laterodorsal and one pair parasagittal); ventral surface of abdominal segments I–VIII with six setae at midlength (two pairs ventrolateral and one pair parasagittal). Legs separated by 8–12 times diameter of coxa.
Etymology. From Tupi-Guarani language; Tuja = adult, mitã = child; allusion to neoteny. Gender: neuter.
Composition and distribution. Only T. plenalatum (Brazil: Minas Gerais).
Tujamita plenalatum sp. nov
Type material. Holotype, male, “Brazil, Minas Gerais state, Itajubá municipality, Municipal Biological Reserve of Serra dos Toledos (22°25′21.3″S 45°22′06.2″W), 1,358 m, malaise trap, 15.X.–8.XI.2015, Rosa S.P. & Dias D. leg.” (deposited at MZUSP). For information on paratypes and other material examined see the Supplementary Text.
Diagnostic description. Adult male (Fig. 5e,f). Labrum four times as wide as long, anterior margin rounded; maxillary palpus 5-segmented; pronotum with lateral carina; elytra not tapered apicad, apices contiguous; hind wing 0.2 times as long as elytra; abdomen relatively wide, with sides rounded and tapered apically; phallus and parameres together 1.1 times as wide as long; endophallus emerging from dorso-apical oval orifice on dorsal surface of phallus. For more details see the generic diagnostic description and the Supplementary Text.
Etymology. From Latin; plenus = full, alatus = winged; allusion to complete elytra.
Jurasaidae fam. nov
(Figs. 2–5, S1–S12).
Type genus. Jurasai gen. nov.
Diagnostic description. Male. Body soft (Fig. 5); head (Figs. S2a, S4a, S5d, S6a–c,f,g, S7a,b,d) declivous, frontoclypeal suture absent, labrum sclerotized, free, separated from head capsule by membrane; antennal insertions elevated and visible from above; antenna filiform, with 11 antennomeres. Mandible falcate, unidentate; maxillary cardo, stipes, galea and lacinia indistinct, last palpomere shorter than all remaining palpomeres combined; gular sutures separated; two posterior tentorial pits present. Thorax (Figs. S2b, S4b–h, S5d, S6e,j,k, S7f–h) with pronotum wider than long, narrower than elytral base; prosternum keel-shaped, strongly convex medially, prosternal process not extending beyond coxae; procoxal cavities open internally and externally, widely separated by prosternal process; mesoventrite evenly sclerotized with anterior margin arcuate, separated from metaventrite by feeble suture; leg (Figs. S1e,g, S5b,e) with meso- and metacoxae oblique, pro- and mesotrochantin exposed; procoxa, mesocoxa and mesal part of metacoxa conical and strongly projecting, tibial spurs long, tarsi 5-5-5, without lamellae or pulvilli. Elytra (Figs. 5, S1a,f, S5a) soft, irregularly punctate, epipleura gradually narrowed posteriorly; wings (Fig. S8) folded longitudinally in resting position, longer than elytra, hiding the abdomen in dorsal view, veins reduced or blurred, cells and transverse cross veins absent, apical field with three triangular sclerotizations and a notch between two most apical ones. Abdomen (Figs. S1h, S4h, S5f) with five free ventrites (i.e., sternites III–VII); sternites II and VIII largely membranous and concealed; sternite IX (Figs. S2e, S4l, S5g, S7m) with apex bilobed, densely pilose and projecting above phallus and parameres; punctures surrounded by usually 2–3 campaniform sensilla in a dog’s-paw pattern (Fig. S1i). Aedeagus (Figs. S2f–h, S4m–o, S5g, S7o–q) trilobate, not entirely retractable into abdomen, phallobase 2.0–2.7 times as long as wide, 1.4–2.6 times as long as parameres, sheathed by a tubular membrane which opens anteriorly into a pair of large balloon-like membranous vesicles; parameres and phallus together 1.1–1.6 times wider than long, basal struts of phallus absent; endophallus emerging from dorso-apical opening, flagellum absent. Female (Fig. 2e,f). Body elongate, wingless, with varying degree of neoteny, but always with at least meso-/metathorax and abdomen larva-like (except for ooporus in posterior margin of sternite VIII) (Fig. S3j,k) (Table S1); leg short, with paired claws; ooporus with pair of membranous lobe-like valves, each with supporting sclerotized plate. Larva (Figs. 1d, 2a–c, S10, S11, S12a–f, i–l). Body cream or milky-white, cylindrical, with few setae; head sclerotized, wedge-like; epicranial sutures and endocarina absent; clypeolabrum triangular, translucent; gula, maxillae and labium fused to each other and to head capsule ventrally, anterior part of labium projected with pair of channels dorsally, fitting apical half of strongly sharpened mandibles, forming a beak-like mouthpart; basal half of mandibles elongate and retracted into anterior 2/3 of head, linked to inner sclerotized rod that extends into prothorax; pronotum with pair of parasagittal sclerotized stripes; prosternum with strongly sclerotized median longitudinal rod; leg short, 5-segmented; trochanter ring-shaped, pretarsus glabrous.
Composition. Jurasai (two species) and Tujamita (monotypic).
Distribution. Brazil (Minas Gerais, Rio de Janeiro).
Phylogenetic analyses
All phylogenetic analyses placed Jurasaidae in Elateroidea and showed an identical backbone topology for the superfamily, with the main clades branching off in the following order: Artematopodidae + Omethidae (incl. Telegeusinae), Throscidae, Eucnemidae, Cerophytidae + Jurasaidae, and the terminal clade of “higher elateroids” sensu Kundrata et al.13 (i.e., Lycidae, Iberobaeniidae, Lampyridae, Cantharidae, Elateridae, Omalisidae, Phengodidae and Rhagophthalmidae). The maximum likelihood (ML) phylogenetic tree of 251 terminals with collapsed branches and Elateroidea families highlighted is given in Fig. 6; the full-resolution tree is in Fig. S13. Jurasaidae were always placed as a sister group of Cerophytum elateroides (Cerophytidae), with 90–95% bootstrap support in the ML analyses and 100% posterior probabilities in the Bayesian (BI) analyses. This clade was sister to the “higher elateroids” with robust statistical support in the BI analyses and weaker support in the ML analyses (Figs. 6, S13). Jurasaidae formed a maximally supported monophylum, with Tujamita sister to a clade formed by two species of Jurasai. We associated the different developmental stages and sexes using molecular markers to confirm the field and laboratory observations (Fig. S14). The uncorrected pairwise genetic distance between cox1 sequences for J. itajubense and T. plenalatum was 28.5% (cox1 was not available for J. digitusdei).
Source: Ecology - nature.com

