The reach-scale manipulation experiment
The reach-scale hydrological manipulation was carried out during August 2017 in a 1.4-km headwater stream located at the upper section of the Krycklan Catchment Study (KCS)28, in northern Sweden (Supplementary Fig. 1). The catchment draining the experimental reach is largely forested, with soils primarily composed of organic-rich deposits in low-lying areas and along the stream29. The experiment was divided in two periods: the drought period (from August 7th to 18th), achieved by damming an upstream lake (Supplementary Fig. 1b), and the background period, which comprised the period before (from August 3rd to 7th) and after (from August 24th to 30th) the drought manipulation (Supplementary Fig. 2). To capture the widest spectrum of responses along the reach, we selected six 50-m segments distributed along the stream (Supplementary Fig. 1b).
At the top and the bottom of the 1.4-km stream (Supplementary Fig. 1b), flumes are installed to estimate hourly discharge (QC5 and QC6, respectively; m3 s−1) based on 10-min water-level observations and stage-discharge rating curves developed from manual discharge measurements55. Hourly stream discharge was estimated every 50 m along the stream using a 2-m digital elevation model as Qi = (UCAi/AC6–C5) × (QC6–QC5), where Qi is stream discharge at channel grid cell i, UCAi is the upslope-contributing area along the stream channel at cell i (m2), and AC6–C5 is the catchment area at C5 subtracted by the catchment area at C6 (m2). The net groundwater inflow to each 50-m grid cell (Qgw,i) was estimated as Qi–Qi–1. Previous studies using hydrologic tracers and hydrometric measurements suggest that this approach provides reasonable estimates of discharge and groundwater inflows along this study reach25,30. We assigned a discharge value for each study segment (QS; m3 s−1) from the modeled estimate. Likewise, we obtained lateral groundwater inputs entering into each 50-m segment (GS; m3 s−1) from the difference between modeled discharge at the top and bottom of the segments.
Further, we obtained the mean stream depth (z; m) and wetted width (w; m) at the six segments (Supplementary Fig. 1b; Supplementary Table 2) from five cross-sectional transects along each segment (cross-sectional measurements every 10 cm). We then combined the segment-specific z and w with the QS data to obtain the mean water velocity (u = Q/z × w; m s−1) for each segment. We derived the stream water residence time (WRT, min) for the six segments at hourly resolution by dividing the segment length (i.e., 50 m) by the mean water velocity. We chose WRT as the hydrological organizer because it correlates with a variety of functional metrics, such as DOM decomposition and chemistry63,64, in-stream metabolism65, hypoxia development66, and nutrient uptake and delivery rates67. Note that estimates of Q and WRT were similar (± 10%; n > 50) to those obtained from salt releases made throughout the experiment period and previous studies25 at different locations along the stream.
Discrete sampling during the experiment
We manually collected surface stream and hyporheic water at the bottom of each segment on five occasions (three during drought and two during background conditions, Supplementary Fig. 2b) to determine the concentrations of major electron acceptors (i.e., dissolved oxygen (O2), nitrate (NO3−), sulfate (SO42−), and carbon dioxide (CO2)), major reduced products (i.e., ammonium (NH4+) and methane (CH4)), dissolved organic carbon (DOC), as well as a set of basic physicochemical parameters (i.e, temperature, pH, and conductivity). Hyporheic samples were collected from 0.6- to 1.5-m-long PVC wells (10-cm Ø, screen length = 10–15 cm) installed in the hyporheic zone (depth = 25–50 cm) using a peristaltic pump. To minimize pumping effects and avoid artificial gas exchange, we pumped slowly and limited our withdrawal of water to a maximum of 250 ml per well. We additionally installed near-stream groundwater wells (depth = 50–100 cm) at the four main groundwater input zones discharging into the stream25 and sampled them using the same methodology described for the hyporheic wells. For each water sample, we measured in situ conductivity, temperature, and O2 concentration with portable meters (YSI, CA, USA). Samples for pH were collected in high-density polyethylene bottles, and filled completely without air bubbles. For DOC, SO42−, NO3−, and NH4+ analysis, samples were filtered (0.45 μm) in the field and collected into clean, pre-rinsed polyethylene bottles. For CO2 and CH4, a separate 5-ml sample of bubble-free water was taken and injected into a 22.5-ml glass vial (containing nitrogen gas at atmospheric pressure) sealed with a rubber septum. The vials were prefilled with 0.5 ml of 0.6% HCl to shift the carbonate equilibrium toward CO2. Samples were kept cold (for pH, DOC, CO2, and CH4) or frozen (for NO3−, NH4+, and SO42−) until laboratory analyses.
High-frequency sampling during the experiment
At the bottom of each study segment, we measured continuous surface and hyporheic water O2 concentration (mg L−1), O2 saturation (%), and temperature (°C) at 10-min intervals during the course of the experiment using MiniDOT loggers (PME, USA). In addition, at four of these six segments (S3–S6, Supplementary Fig. 1b), we also measured dissolved concentrations of CO2 at the same frequency with a Vaisala GMT220 sensor (Vaisala, Finland) covered with a highly permeable membrane to dissolved gases but not to water68 and connected to CR1000 data loggers (Campbell Scientific, Canada). Hyporheic sensors were placed in the same wells where the low-frequency sampling of stream hyporheic water was performed (see previous section). We used the continuous O2 data to both validate low-frequency discrete O2 observations and to model stream metabolism (see below and in Supplementary Methods 1).
Network-scale monitoring
To cover the widest spectrum of drainage sizes in the KCS network, we addressed chemical patterns in ten streams ranging from channel order 1 to 5 (sub-catchment drainage area from 0.04 to 68.9 km2; circles with numbers in Supplementary Fig. 1a; Supplementary Table 1). Similarly, to cover a wide range of environmental conditions, we selected streams that drain distinct land covers that are representative of northern boreal landscapes, including forests, mires, and lakes (Supplementary Table 1). For the analyses, we grouped these ten streams into those draining headwater catchments (i.e., stream order 1 or 2; catchment area <1.5 km2; n = 5; Supplementary Table 1) and those that do not (i.e., stream order > 2; catchment area > 1.5 km2; n = 5; Supplementary Table 1). Thus, we used Strahler stream order to categorize these sites. Strahler stream order correlates with a variety of geomorphological metrics, including catchment drainage area69 or stream width70, and is thus a useful organizer for assessing patterns at the network scale71.
At the ten monitoring stations and during two consecutive summers (2017 and 2018), we measured surface water O2 concentration, O2 saturation, and temperature at 10-min intervals with mniDOT loggers (PME, USA), and manually sampled for CO2, CH4, and DOC monthly (during winter) and every second week (during summer and fall). In total, low-frequency chemistry data used for the analysis of the period between 2017 and 2018 derived from ~30 sampling occasions at each stream. In addition, to increase the spatial coverage of high-frequency O2 data, we also deployed O2 sensors in six different locations (four headwater catchments and two larger catchments; circles without number in Supplementary Fig. 1) during the two consecutive summers (2017 and 2018). Similarly, to increase the spatial resolution of low-frequency chemistry data (i.e., O2, CO2, and CH4), we also carried out three synoptic surveys at 22 headwater streams of the KCS during the summer 2018 severe natural drought (triangles in Supplementary Fig. 1a).
Long-term monitoring
Apart from the 2017–2018 network monitoring, we also compiled monitoring data for CO2 and CH4 with a suite of additional chemical and physical parameters for ~9 consecutive years (2010–2018) at the same ten stream-monitoring stations in the KCS (square symbols in Supplementary Fig. 1a; Supplementary Table 1). Long-term monitoring samples were collected monthly during winter and every second week during summer and fall. This time period includes two consecutive summers (2017 and 2018) when sensor O2 data were also recorded. We separated the summer period from the bulk long-term series based on historical (1980–2008) seasonal records in the catchment42. In total, noncontinuous chemistry data used for the analysis of the period between 2010 and 2018 derived from ~100 sampling occasions at each stream. In addition, at each of the ten monitoring stations, we measured Q at hourly intervals using a permanent H-flume55. To normalize and compare Q from the studied streams with different catchment areas, we report specific discharge (mm day−1). To isolate drought hydrological conditions from the rest of the periods, we delineated the specific discharge and grouped studied responses based on percentile distributions of the historical (1980–2018) discharge records in the catchment32: drought (0th–10th percentile; n = 59), low flow (10th–20th percentile; n = 22), baseflow (20th–50th percentile; n = 90), and high flow (50th–100th percentile; n = 193).
Laboratory analysis
pH was measured using an Orion 9272 pH meter equipped with a Ross 8102 low-conductivity combination electrode with gentle stirring at ambient temperature (20 °C). DOC was analyzed by combustion using a Shimadzu TOC-VPCH (Shimadzu, Kyoto, Japan) following acidification to remove inorganic carbon. NH4+ and NO3− were analyzed following the methods G-171-96 Rev.12 and Method G-384-08 Rev.2, respectively, with a SEAL Analytical AutoAnalyzer 3 (SEAL Analytical, WI, USA). SO42− was analyzed by liquid chromatography using a Metrohm IC Net 2.3 (Herisau, Switzerland). Finally, the concentration of CO2 and CH4 in the headspace gas samples was determined using a GC-FID Perkin-Elmer Clarus 500 (Waltham, MA, USA) equipped with a methanizer operating at 250 °C and connected to an autosampler Perkin-Elmer Turbo Matrix 110 (Waltham, MA, USA). Concentrations of other species of the DIC system (i.e., HCO3− and CO32−) were also determined using the stream pH, equations for carbonate equilibrium, and Henry’s Law72. Free dissolved CO2 was the predominant DIC form, accounting for >95% of DIC. Accordingly, HCO3− and CO32− were discarded from the analysis due to their minor contribution to the overall DIC composition.
Data treatment and statistical analyses
For each study segment and experiment day, we used continuous O2 measurements to estimate gross primary production (GPP) and ecosystem respiration (ER) with the open-channel single-station method73. We used Bayesian inverse modeling to estimate both GPP and ER74,75. A more detailed description of the stream metabolism modeling, quality assessment, and potential uncertainties can be found in Supplementary Methods. Note that here we only focused on ER, which is an integrative estimate of the ecosystem aerobic respiration occurring in the stream. We compared ER rates between drought and background periods using a nonparametric Wilcoxon Signed-Rank test. Further, we evaluated the relationship between ER and stream WRT to test the effects of drought on in-stream aerobic respiration.
To explore whether drought influenced the overall distribution of redox-sensitive solutes and gases in the stream, we built a principal component analysis (PCA) with the surface and hyporheic water O2, CH4, SO42−, NO3−, and NH4+ concentrations from the samples collected during the experiment. We evaluated the dependency of the resulting scores of the PC1 (dependant variable) on stream WRT (independent variable) using linear and nonlinear regression models. We selected and reported the model with a higher coefficient of determination (r2). Differences in the distribution of surface water PC1 scores between background and drought conditions were visually inspected with Kernel density plots and statistically tested using the nonparametric Wilcoxon Signed-Rank test.
We additionally assessed specific molar ratios of reduced to oxidized chemical forms (i.e., NH4+:NO3− and CH4:O2). The NH4+:NO3− ratio provides insight into the potential redox-driven changes in nitrogen cycling. Accordingly, an accumulation of nitrogen as NH4+ rather than NO3− represents constraints on nitrification and increasing rates of denitrification34. Demand for NO3− under reducing conditions is very high, as NO3− is the most energetically favorable electron acceptor in the absence of oxygen34. We also used the CH4:O2 ratio to provide a synthesis of the full range of terminal electron-accepting processes in the sample. Low values of the CH4:O2 ratio indicate that aerobic pathways dominate the metabolic balance, while increases of CH4:O2 ratios represent a shift toward a dominance of anaerobic over aerobic metabolic processes36,37. To examine whether drought drove similar redox responses under experimental and natural conditions, we compared the relationship between stream WRT and surface water CH4:O2 molar ratios along the experimental stream reach during summers 2017 (experimental drought) and 2018 (severe natural drought).
To explore the influence of drought on the stream metabolic balance during the experiment, we compared molar deviations of O2 and CO2 from atmospheric equilibrium (ΔO2 and ΔCO2, respectively) for the discrete and high-frequency observations. The stoichiometry between O2 and CO2, in aquatic ecosystems is of particular interest because it provides insight into the dominance of the different metabolic pathways involved in the production and consumption of organic matter76. For instance, aerobic respiration of organic matter normally leads to ΔO2 and ΔCO2 relationships falling around the 1:–1 line. Deviations from this stoichiometry can be attributed to nonbiological processes (i.e., interactions of CO2 with the carbonate system77) or to anaerobic respiratory pathways that produce CO2 and CH4 without consuming O278. We calculated ΔO2 and ΔCO2 from differences between the measured aqueous concentration of the gas (Cw) and its concentration in equilibrium with the atmosphere (Ca). Equilibrium concentrations were calculated from temperature and barometric pressure72. Different statistical analyses on ΔCO2:ΔO2 observations were used to test the treatment effects on the central tendency and dispersion of these data36.
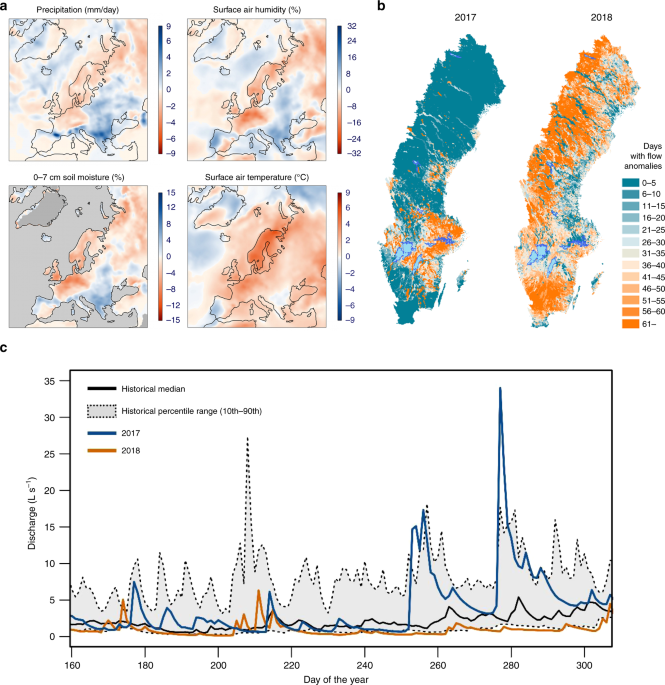
To assess the network-scale effects of the 2018 severe drought on the stream surface O2 availability, we compared the 10-min O2 saturation dynamics at the 16 stream-monitoring stations during 2017 and 2018. We evaluate the effect of catchment size (as stream order) on stream surface O2 availability using nonparametric 10th, 50th, and 90th percentile regression. For this, each relationship was computed and plotted as a representation of the central tendency and dispersion of all the data. In addition, we explored patterns for the molar CH4:CO2 ratio during 2017 and 2018 to quantify the extent to which drought induced methanogenesis. Given that the quantification of whole-stream anaerobic metabolism is more difficult than aerobic respiration as a routine part of metabolism studies35, molar ratios between CH4 and CO2 have been proposed as an indicator of methanogenesis in aquatic ecosystems35,49. Although this approach only provides a proxy of process rates, the use of ratios instead of absolute concentrations allows us to isolate the effect of physical processes affecting the absolute concentration of gases (e.g., hydrological mixing or atmospheric reaeration). Finally, because CO2 and CH4 measurements are currently more common than O2 measurements in Swedish monitoring programs, using this ratio allowed us to more broadly assess the influence of drought on stream biogeochemistry.
To explore whether the summer 2018 stream biogeochemical responses extended to past low- flow periods, we analyzed the relationship between specific discharge and the molar CH4:CO2 ratio at the surface water of five headwater streams draining contrasted boreal catchments during summer for the period compressed between January 2010 and October 2018. Nonparametric 10th, 50th, and 90th percentile regression for all sites was computed and plotted as a representation of the central tendency and dispersion of the data. Specific site-to-site responses to different discharge levels were assessed with the locally weighted regression model (Loess). Differences in the distribution of surface water CH4:CO2 ratio across contrasting discharge conditions were visually inspected with Kernel density plots and statistically tested using the nonparametric Wilcoxon Signed-Rank test. Finally, to examine the significance of drought in promoting anaerobic respiratory processes at a wider regional level, we compared the CH4:CO2 ratio in the KCS during drought with the CH4:CO2 ratio of seven additional headwater catchments in Northern Sweden sampled before the summer 2018 severe drought and used as a reference for nondrought conditions (Supplementary Fig. 1a).
All statistical analyses were conducted with the R statistical environment (R Core Team 2018), except for PCA analysis, which was done with the software XLSAT (XLSTAT 2019.1, Addinsoft SRAL, Germany). In R, we used the packages “stats”, “nlme”, and “vegan” to calculate and visualize linear and nonlinear regression models as well as nonparametric Wilcoxon Signed-Rank tests. We also used the “quantreg.nonpar” package to compute and visualize nonparametric percentile regressions. Statistical tests were considered significant when p < 0.05.
Source: Ecology - nature.com
