Saline alkaline (soda) lakes are extreme environments for most organisms. Although microbial extremophiles often flourish in these settings, few animals survive under the prolonged stressed conditions that characterise these lakes7,24,47. Hydrologically closed lakes nonetheless are dynamic environments that can undergo rapid, and sometimes extreme, changes in salinity and alkalinity during periods of freshening or evaporation that are usually tied to increased dilute inflow from the drainage basin (rivers and direct rainfall), varying recharge from groundwater, or loss of inflow during arid periods64. Such changes are often linked to climate variations on annual to decadal to centennial or millennial timescales, but also to longer-term tectonic modifications of the hydrology (e.g., river diversions) and hydrogeology in the evolving rift65,66.
Hypersaline lakes can become brackish or even dilute within a few years of freshwater inflow. Lake Bogoria, for example, a perennial highly saline (>50 g/L TDS in summer 2006) lake in the central Kenya Rift, became fresh enough to support crocodiles, fish and hippopotamuses a few years later after prolonged heavy rains67. In saline lakes, the biological diversity often increases initially near fluctuating lake margins where local littoral and shallow-offshore freshening accompanies rising lake level, while low diversity persists in deeper parts of the lake because of ponding of dense saline water in lake-floor depressions or chemical stratification in the water column68. Together these often lead to anoxia in the hypolimnion or monimolimnion. In contrast, carbonate and evaporite precipitation typifies falling and lowstand conditions in shallow ephemeral saline lakes4. Saline pans and residual pools, often near spring-fed shrinking evaporitic lakes, are stressful environments that typically preserve few biogenic structures2. Hypersalinity and oxygen-depletion can result from water stratification during oligohaline (brackish) phases (e.g., flooding by fluvial inflow), increasing stress on benthic organisms. The presence of macroburrows in magadiite, which precipitated in the most saline of the large modern lakes in the East African Rift, is therefore surprising. A critical issue is the chemical environment of the submerged or briefly exposed lake floor during burrowing.
Magadiite, first described at Lake Magadi18,19,20,33, is the most common of several rare sodium-silicate minerals (including kenyaite, makatite and kanemite) that form in saline, highly alkaline lakes69,70,71,72,73. Magadiite at Lake Magadi is present in brown and dark green lacustrine muds as irregular nodules, up to ~15 cm long and 5 cm thick, thin (<5 mm) lenses and dispersed patches (mm–cm scale), and as thin laterally-continuous laminae (<3 mm) and beds (<15 cm) interlayered with zeolitic lacustrine silt and mud, some rich in dark brown to black organic matter18,19.
Chemical conditions for magadiite formation are well known, and the mineral can be synthesized in the laboratory74,75,76,77,78. It has been proposed that bedded magadiite interlayered with muddy sediments formed at the chemocline of a stratified lake (intermittent stratification or meromixis)16,18,19,20. In that model, the mineral precipitated where evaporated highly alkaline, silica-rich sodic brine became overlain by more dilute, less dense water, as for example during seasonal flooding by runoff. The lowered pH at the fluid interface would decrease silica solubility in the underlying brine16,17,18,19,20. Magadiite forming in the water column would then settle gravitationally on the lake floor, forming a soft, possibly gelatinous layer composed of fine spherulitic grains21. In a stratified water column, the different densities of the water masses would limit wind mixing or overturn until evaporation of the surface waters (epilimnion or mixolimnion) led to approximately equal densities. For pure magadiite to form implies that little detrital sediment was then present in the surface waters. Lowering the pH in a sodic brine could also have been induced by biogenic CO2 released from decay of degrading organic matter19, or possibly from pulses of CO2 of geothermal (upper mantle to lower crustal) origin79. One cm of magadiite is thought to form in about 20 years19.
Modern examples of magadiite precipitation are rare, so the physical state of magadiite soon after precipitation is unclear. However, recent magadiite formation has been documented in a core from Lake Kitagata (Uganda), a small shallow (9 m deep) perennial saline (>170 g/LTDS) alkaline (pH 9.7) crater lake fed mainly by groundwater80. There, the magadiite forms in the silica-rich (256 ppm SiO2) water column as a primary mineral that settles upon the substrate and in places forms thin beds. Some magadiite grains contain muddy inclusions implying some in situ formation by intrasediment growth. The lake is meromictic with a chemocline at 1 m depth81.
In contrast, the formation of magadiite in an evaporative brine pond has been reported at Alkali Lake, Oregon, USA82. That pond was fed partly by dilute alkaline well-water. The authors reported “ball-like aggregates of white semi-crystalline material with patchy opalescence”. X-ray diffraction analyses confirmed magadiite and clay minerals. Magadiite might also precipitate directly from solution by evaporative concentration20, as is the case of littoral muddy plains at Lake Magadi that were temporarily covered with shallow brine undergoing evaporation21. The thickest magadiite unit at the base of the Lake Kitagata core is crumbly, overlies a palaeosol, and probably formed by capillary evaporation when the crater lake had almost desiccated81. Magadiite has also been reported from Late Pleistocene and recent sediments in Lake Bogoria64,72,83,84, Lake Chad70,71,73, Manga in Niger71, Sua Pan in Botswana85, and Malha crater lake in Sudan86.
The ichnologic evidence provides clues to the magadiite origin at Lake Magadi. Whether magadiite precipitated by evaporation of a shallow, silica-rich sodium carbonate brine or in a deeper chemically stratified alkaline lake, the lake-floor environment during colonization – benthic or temporarily subaerial – would initially have been hostile to most organisms except bacteria and archaea. The contact between the trace-bearing magadiite bed and the overlying lacustrine silt and mud is thus regarded as the primary colonization surface, although some burrows penetrated the magadiite from up to ~8 cm above the contact. In some examples, the burrow fill is coarser-grained than the overlying silt or mud, and represents a period of time on the colonization surface where these sediments were locally available before continued mud deposition. Accordingly, environmental conditions during colonization were either those associated with the initial silt or mud deposition upon the underlying magadiite substrate, or from this colonization surface during a short depositional hiatus.
The exposed sediments with distinct magadiite beds, including the one containing burrows, show repeating facies successions (‘cycles’) on a cm–dm scale that give clues to the pattern of sedimentation when the ichnofossils were formed (Fig. 4). We interpret each cycle to begin with lacustrine silt and mud and terminate with bedded magadiite of variable purity and thickness. Each magadiite bed is overlain by dark to greenish brown zeolitic silt, fine sand or mud (Facies a), some of which shows indistinct lamination. The contact, interpreted as a small-scale flooding surface, is usually sharp, but locally irregular because of minor compaction of muds into the underlying soft magadiite, local W-E slippage from the Magadi horst, or local erosion. These silt, fine sand and mud layers, in turn, are commonly succeeded by a few cm of interlaminated mud and magadiite (Facies b), which is commonly undulating or wavy in sectional view (Fig. 2c). Those laminae are continuous in some units (Fig. 2d) but broken or disrupted with small-scale disconformities in others (Fig. 2e). Where undisturbed, the laminae lie parallel to laminae in the underlying brown mud and silt if present, and commonly inherit underlying morphological irregularities in the substrate. These laminated units are then capped either by white or grey magadiite (Facies c) or magadiite containing thin brown mud laminae (Facies cm), typically with sharp upper and lower contacts (Fig. 4). The thickest magadiite beds have fewest impurities. Some are streaky in outcrop with eastward-dipping magadiite layers thinning westwards from the horst (Fig. 2b).

Field photographs that show repeating (cyclic) patterns in sedimentation at the study site. Each arrow indicates one shallowing-upward cycle. Facies codes a–d are described in the text. Photographs taken by RWR.
The critical question in interpreting these cycles is whether the magadiite bed(s) records precipitation in a shallow lake where magadiite formed by (1) evaporative concentration of a silica-rich sodic brine in shallow oxygenated water, or in (2) a deeper stratified lake with precipitation at a chemocline (Fig. 5). The burrows imply at least partial or temporary oxygenation of the lake floor when they formed, and (or) a brief period of exposure before continued sedimentation. That in turn has implications for understanding the origin of bedded magadiite.
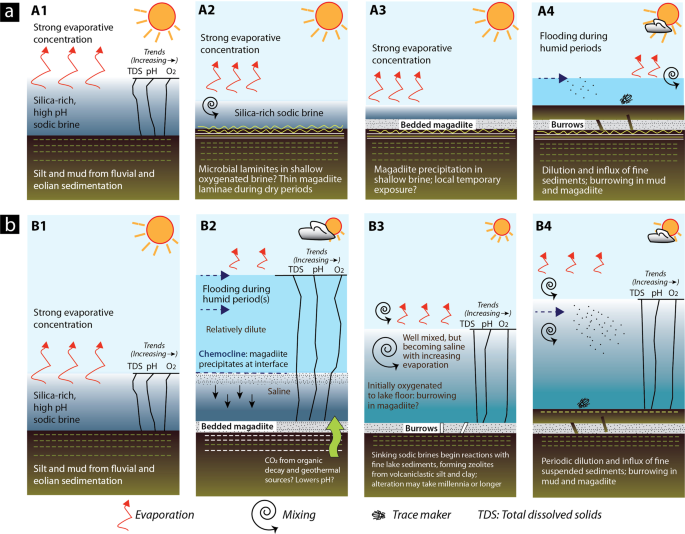
Models for the origin of bedded magadiite and its trace fossils. (a) A1 to A4 show the shallow-water model, which is best supported by our evidence. A1. Following deposition of fine clastics, lake water evaporates producing an alkaline sodic brine with high pH (>10). High aqueous silica concentration. A2. The brine attains saturation with respect to magadiite which precipitates seasonally, alternating with siliciclastic mud in shallow (<1–5 m?) water to form mm-scale laminites. Undisturbed laminae imply bottom-water anoxia. Where the lake floor is shallower and at least temporarily oxygenated, microbial mats might influence or control benthic sedimentation. A3. With continued evaporation the shallowing brine becomes supersaturated with respect to magadiite, which precipitates and settles rapidly as soft gelatinous material that extends laterally for hundreds of metres and forms thinly bedded magadiite on the lake floor. Some beds become contaminated by siliciclastic sediments derived from ephemeral-stream inflow (facies cm) and eolian dust (or volcanic ash). In lake marginal settings and periods when lake level is unusually low, some shallow magadiite might become subaerially exposed. This provides an opportunity for organisms (subaerial and shallow subaqueous: aerobic or dysaerobic that need oxygen) to traverse the littoral zones and feed on newly exposed microbial organic matter. A4. Rise in lake level delivers fine siliciclastics and provides fresher water. Organisms adapted to fresher waters burrow into underlying siliciclastic sediments and magadiite. Some open burrows become filled by fine sediments or remain open for minerals precipitated during later evaporation. (b) B1 to B4 show the perennial lake model in which magadiite precipitates at the interface of a stratified lake, when shallower dilute lake waters move across a denser sodic brine. After deposition of fine clastics at the interface, lake water evaporates producing a sodic brine with high pH. Aqueous silica concentration remains very high. B2. Flooding linked to increased rainfall (or drainage diversion?) dilutes Lake Magadi and a stratified lake develops. Magadiite then precipitates at the chemocline. Initially the magadiite may alternate with mud, forming laminites. B3. With evaporation and shallowing the waters become mixed and oxygenated to the lake floor. Burrowing occurs while conditions remain favourable. B4. Flooding introduces fresher water and sediment. Evaporation gradually increases the salinity. Drawing produced by RWR using Adobe Illustrator CC v. 23.0.4.
The cycles (Facies a to c) are interpreted as shallowing-upward successions in shallow (dm to a few metres?) water (Fig. 5 A1–A4). The magadiite beds, which represent the most saline part of the cycle with high Na+ and aqueous SiO2, are periodically flooded by runoff that introduces detrital fine sand, silt and clay that are deposited upon a magadiite substrate. The aqueous environment during flooding was probably oxidizing, but initially brackish given that the inflow water flooded saline water and (or) a saline substrate. Mud-silt sedimentation continued from seasonal runoff but as the water shallowed and increased in salinity, it became sufficiently saline for magadiite precipitation, which for a period alternated with thin mud laminae. Some alternations are regular (Fig. 2d), implying that they could have been seasonal with magadiite precipitating in the drier periods and mud settling gravitationally during the rainy seasons. This speculation is unconfirmed. These couplets, however, resemble microbial laminites in lacustrine upward-shallowing cycles. Most laminae are parallel and unfractured, implying that they formed and were preserved in shallow water. Some that are broken or crinkled may record brief periods of subaerial exposure or near exposure (Fig. 2e). A darker layer <1 cm thick that locally underlies the laminite zone (Fig. 4) could represent a former layer of bacterial reduction below organic laminites. A thin dark microbial (?) lamina is also present below some magadiite beds (Fig. 2e).
The magadiite beds that cap the cycles represent the most saline (Na- and silica-rich) fluids. The thickest and purest beds might have formed in shallow brine upon underlying subaqueous laminites. The thinner and more irregular magadiite layers could have been disrupted by intermittent subaerial exposure or intrasediment growth at a very shallow water table, but the overlying siliciclastic mud and silt shows no clear evidence of exposure. Were the thicker magadiite beds subaerially exposed? Where subaerially exposed today by erosion, soft plastic magadiite rapidly becomes encrusted with efflorescent trona, and soon forms a thin (1–3 mm) hard grey crust. The uppermost surface (bedding plane) initially develops small (2–4 mm wide) cracks on exposure and begins to transform to quartzose chert with reticulate crack networks during early diagenesis (see below). This was not observed at the study site because the bedded magadiite was buried until excavated but has been observed elsewhere at several locations in the southern Magadi basin. Contemporary development of a hard silica crust, resulting from dehydration during exposure, would have prevented, or at least inhibited, the tracemakers from burrowing in still soft, plastic magadiite. The sharp, locally irregular upper contact also implies that the magadiite remained submerged during the next flooding event that introduced siliciclastic mud. Magadiite intraclasts were not found in the overlying basal muds. The most likely setting for this scenario would have been in a shallow (<1 m deep at the study site?) lake and laterally adjacent mudflats.
The alternative explanation would be for magadiite precipitation in a stratified lake16,19,20. Cycles in a stratified lake would result from mud-silt deposition during relatively humid phases followed by evaporation in a lacustrine offshore setting (Fig. 5B1–B5). After precipitation of magadiite in a shallow silica-rich alkaline brine, inflow of freshwater bearing fine clastic sediment might lead to temporary stratification, with magadiite precipitating in the water column at the fluid interface, where the pH was lowered along the contact of the upper and lower fluids. Alternatively, when CO2 was being released from biogenic or deeper geothermal sources into the bottom waters (hypolimnion or monimolimnion). Clastic laminae would form during seasonal inflow of mud and silt from ephemeral streams similar to the lake-shallowing model, but without photosynthetic benthic microbial mats because of anoxicity below the fluid interface (i.e. chemocline). Those inflows would likely form hypopycnal plumes, as happens today at Lake Bogoria87. Evaporation would then lead to precipitation of thicker magadiite beds in the offshore brine until aqueous Si and Na+ declined in the surface waters. Further evaporation would lead to trona precipitation, but this is not evident in the exposed HMB sections. Decreasing pH, resulting from organic matter decay or geothermal inflow, would not necessarily be seasonal, making this a less likely option for producing repeating couplets observed at the study site and elsewhere. The ichnologic evidence is less consistent with this scenario because it implies that the producers were able to migrate deeper into the water body. Assuming that the producers might have had similar environmental range to the beetles in modern Lake Magadi, this scenario is unlikely, however, because the latter live either in very shallow pools (i.e. Coelostoma tina) or are essentially terrestrial (i.e. Lophyra pseudodistans)38,39. The lake floor at neighbouring Nasikie Engida, a small shallow (1.6 m deep) saline alkaline lake northwest of Lake Magadi (Fig. 1b), was anoxic at only 1 m water depth when measured88. Although tiger beetle larvae and staphylinids can survive periods of flooding and even anoxia55,56,63, they inhabit marginal areas that are often subaerially exposed. An offshore setting in a stratified lake would be flooded perennially over time periods much longer than those during which burrowers could survive.
After deposition of the magadiite beds capping the exposed lacustrine sediments (Fig. 4), the lake expanded with deposition of black and dark brown, locally organic-rich mud preserving fossil fish. This basinwide ‘event’ of unknown duration is recorded along the N-S basin axis because this fish bed is a marker horizon in outcrops north and south of Lake Magadi19,27, implying deeper stratified waters with an anoxic monimolimnion. This fish-bed marker was identified as far north as southeastern Nasikie Engida. Magadiite in black anoxic muds has also been recorded at Lake Bogoria71,72,83,89, showing that the stratified-lake model for bedded magadiite genesis is feasible but modern examples of bedded magadiite are unconfirmed. That model does not, however, concur with the sedimentological and ichnologic evidence for shallow waters and lake-floor oxygenation during sedimentation at the Magadi study site.
The Pleistocene Green Beds, which underlie the HMB disconformably, show a similar regressive succession in outcrops south of Lake Magadi27, where oxidized lacustrine silt and mud, some with large (cm-scale) burrows (Fig. 6), pass upwards into thinly bedded cherts in ‘cycles’ with a similar (cm-to-dm) scale to those of the HMB40. Bedded cherts in the Green Beds show possible stromatolitic layering, burrows, salt crystal pseudomorphs (calcite and Na-carbonates?), tepee and petee structures, and other features that imply shallow-water sedimentation on playa floors or upon broad littoral mudflats19,21,22,32,41. Most of those cherts, however, lack the characteristic reticulate crack-patterns commonly associated with diagenesis of a magadiite precursor19,90. Those patterns alone, however, are not necessarily diagnostic of former magadiite. The Green Beds chert precursor(s) might have been a siliceous gel (which form today at Nasikie Engida91, Fig. 1b), opaline silica, a carbonate mineral21, magadiite, or an as-yet-unidentified phase.

Burrows in the Pleistocene Green Beds from the southern end of Lake Magadi (see Fig. 5 in ref. 27) that are very similar to those in the HMB. (a) Burrows originate at the contact between lacustrine muds and an overlying darker layer of mud. The thin (1 mm) discontinuous white laminae below the contact are silica. Some of those patterns on bedding planes resemble those in Fig. 3F, but their nature remains uncertain. (b) Close-up of a single burrow showing infill contrasting with the host sediment. Photographs taken by RWR.
Magadiite, a metastable hydrous mineral, is a precursor of some lacustrine cherts. During early diagenesis, magadiite undergoes physical and chemical changes including loss of water and Na, and early recrystallization. Soft magadiite transforms progressively to microcrystalline quartz. Early diagenesis includes leaching of Na by percolating water followed by rapid recrystallization18,19. Using stable isotopes of oxygen at Olduvai (Tanzania), it has been shown that magadiite could ‘spontaneously’ recrystallize to quartz in saturated sodic brines20,92. This transformation sometimes involved intermediate phases including kenyaite [Na2Si22O41(OH)8•6(H2O)]19 and moganite (SiO2)93.
The timing of these diagenetic fluid-mineral reactions remains unclear but can be rapid (<10 ka and probably much less) because some magadiite in the High Magadi Beds has altered to kenyaite, moganite and chalcedonic microcrystalline quartz18,19,33,93. Early and rapid diagenesis also has implications for the interpretation of traces that were originally produced in semi-soft magadiite: first, their dimensions might change, reflecting shrinkage or expansion52 but early lithification could also reduce compaction. Second, uncemented siliciclastic fills might be physically flushed during or soon after lithification, especially with subaerial exposure in a closed lake with fluctuating level, leaving open pores or providing pore space for cementation by other minerals such as calcite or silica. The sharp burrow margins in the examples illustrated here indicate a semi-firm, but still moist, substrate.
Based on the evidence and discussion above, we infer that the fine sediments overlying the magadiite described in this study, although later altered to zeolites during early diagenesis33,37, record a period of more dilute lake water after the main phase of magadiite formation. During that time of generally fresher conditions, the trace makers inhabited the shallow lake floor, that for short times might have been subaerially exposed, and burrowed into the soft magadiite substrate. Continuing freshening is implied by the fossil fish upwards in the succession, notably in a dark, parallel-laminated layer approximately 10 cm above the top of the bioturbated magadiite bed. This cichlid-bearing layer has been radiocarbon dated at ~9,100 y BP94,95,96 placing it within the African Humid Period.
Chemically stratified lakes typically develop anoxic bottom-waters – a lacustrine environment unfavourable for macroscale trace-makers. Chironomids can tolerate extreme dysoxic conditions in lacustrine environments45,50,97. In contrast, most other burrowers, including water-scavenger beetles, which are possible producers of the large burrows at the study site, cannot survive extreme or prolonged dysoxia. Nevertheless, tiger beetle larvae, staphylinid beetles, and heterocerid beetles ― all known from hypersaline lake margins in the Kenya Rift ― are able to survive periods of flooding, and tiger beetles can even tolerate anoxic conditions during flooding for days to months55,56,63. In addition to freshening, increased dissolved oxygen might have developed on the lake floor. That would imply that the trace makers burrowed into an older magadiite bed rather than being contemporary with its precipitation, or they formed the burrows soon afterwards. Integration of ichnologic and sedimentological evidence implies that changing environmental conditions, whether freshening or brief subaerial exposure, might have favoured rapid colonization in a previously harsh benthic setting.
Lake water depth during magadiite sedimentation is unclear. A maximum elevation of 656 m for the HMB shoreline based on stromatolitic carbonates that extend southwards into Tanzania has been reported98 but remains unconfirmed by other sedimentological evidence21,27. A palaeoshoreline with stromatolite-coated gravel ~12 m above Lake Magadi along the fault scarp directly east of the Magadi horst27,28, and at many localities in the axial graben, provides clear evidence of a former HMB lake level that might have been contemporary with sedimentation at the study site. That lake would have been saline and alkaline during magadiite sedimentation and perhaps only a few metres deep at its maximum depth by analogy with other examples64 (e.g., Lake Bogoria89).
The laminated and muddy facies that overlie the magadiite beds record a change to deeper water above a more extensive flooding surface and by implication, more dilute and oxic conditions, at least in the waters that extended downwards to the lake floor during periods of burrowing. Interlaminated magadiite and muds, which overlie the burrowed magadiite bed, might indicate seasonal or irregular stratification of the water column with magadiite particles settling by gravity. The fossil fish record an expanded lake across much of the axial graben, implying deeper fresher water after a phase of more saline conditions.
Evaporitic soda lakes host simple food webs7,47. Modern Lake Magadi has a highly productive trophic web dominated by the heterotrophic bacteria Arthrospira sp., with detritus feeders and omnivorous chironomid larvae and copepods, the fish Alcolapia grahami, and several birds, including lesser flamingos, as primary consumers7,24,47. The addition of large burrowing insects to the aquatic benthos, most likely scavenging or predator beetles, would imply added complexity to the Lake Magadi food-web during periods of decreasing hydrochemical stress. Staphylinid beetles, however, are also primary consumers that feed on microbe-rich sediments at the sediment-air-water interface, and are well known from hypersaline lacustrine settings61,99. They can also survive inundation within their bottle-neck burrows, feeding on microbes during subsequent brief exposure-events60,63.
Although integration of ichnologic and sedimentological evidence is common practice in facies analysis, this process implies linking datasets that typically have marked differences in terms of temporal resolution and implications. Benthic colonization by burrowers may reflect short-term changes in environmental conditions that do not leave a physical or chemical record in stratigraphic successions. In this example, colonization in magadiite might have resulted from freshening and oxygenation of a shallow lake bottom and (or) brief exposure of the siliceous substrate. The bioturbation is decoupled from magadiite formation in terms of physical and chemical conditions. Detailed ichnologic analysis provides the clues for changing environmental conditions that are otherwise unrecorded or unrecognised in the sediment record.
Source: Ecology - nature.com

