Land-use and land-cover changes have impacted many natural ecosystems to provide ecosystem goods and services for an ever-growing human population1. This often has unintended consequences that may threaten biodiversity and ecosystem health2. Such consequences include changes in local, regional, and global climate3, alteration of natural habitats4, pollution of land, air, and water5,6,7, and changes in landscape structure, i.e., the total area and spatial configuration of ecosystems8.
Natural ecosystems offer habitat for many species, and human landscape modification typically involves habitat loss as well as the breaking up of continuous habitats into smaller remnant patches (fragmentation)9,10. Consequently, landscapes may lose connectivity, i.e., the degree to which they facilitate movement of organisms and their genes among patches11,12. Landscape connectivity can be quantified in three ways: structural landscape connectivity, potential functional connectivity, and actual functional connectivity13. Structural landscape connectivity can be determined from physical attributes, based on maps alone without reference to organismal movement behaviour. Potential functional connectivity relies on a set of assumptions on organismal movement behaviour to implement an organism perspective, e.g. by mapping a species’ habitat and setting a dispersal threshold. In contrast, actual functional connectivity refers to observed data (e.g., patch occupancy, radio tracking, mark-recapture, or molecular genetic data) that reflect actual rates of the exchange of individuals (or their genes) and may be used to test models of structural or potential functional connectivity13. As landscape connectivity facilitates organism dispersal, gene flow, and many other ecological functions of a landscape14, its erosion is a major concern for wildlife population survival, due to increase of extinction risk, loss of species diversity, and disruption of major ecosystem services11,15,16,17. For these reasons, connectivity is considered a key aspect of land-use planning and conservation management18,19.
Connectivity maps are sensitive to the way connectivity models are conceptualized and implemented20,21, and there is no general consensus on which approach should be used preferentially to support planning. The most common approach to modelling connectivity is the focal species approach10,22. This bottom-up approach considers one or a few species that serve as surrogates to characterize connectivity for a larger suite of species23. It aims to evaluate the potential functional connectivity of a species’ habitat by taking into account species-specific dispersal thresholds and modelling the impeding effect of different feature types on movement as resistance values24. However, because species differ, e.g., in habitat requirements, body size, dispersal ability, or lifespan, the effect of habitat fragmentation on functional connectivity can differ importantly25,26. For this reason, the effectiveness of such an approach to evaluate connectivity for a suite of species is highly debated27,28,29. In addition, large-scale analyses may require large numbers of focal species to represent diverse habitat types20. To overcome these limitations, there is growing interest in applying a top-down approach, which does not rely on biological or ecological characteristics for specific taxa to model large-scale connectivity maps for management and planning efforts30,31, recently known as a “species-agnostic” approach (e.g.32). More specifically, these models are solely based on quantifying the degree of unnaturalness in the landscapes which are caused by human modifications29 as well as other ecological and geochemical processes. Indeed, the approach is based on the assumption that natural terrestrial areas facilitate connectivity, and the higher the degree of human modification, the more the ‘ecological flow’ is restricted. Models based on structural connectivity are not new. For instance, effective mesh size30 likewise does not make reference to species characteristics. It is important to note that even a species-agnostic approach will be parameterized with a certain group of species in mind, in this case terrestrial organisms20,33.
Because large water bodies, despite being natural features, may disrupt the movement of many terrestrial organisms, some authors have treated them as a barrier34. The hydrological connectivity of aquatic habitats, on the other hand, should be modelled separately as it has a linear network structure.
More generally, focal-species and species-agnostic approaches both are known to be sensitive to resistance values, which are often based on expert opinion, and a rigorous assessment of the sensitivity to parameter settings is required35,36,37. For instance, Koen et al.28 successfully validated a species-agnostic model of landscape connectivity with road mortality data of reptiles and amphibians and with molecular genetic data for a mammalian species. Connectivity models are also known to be sensitive to the degree of contrast between high-resistance and low-resistance landscape features, which can be modified with a scaling function38,39. Moreover, Arponen et al.40 demonstrated that the spatial resolution of large-scale connectivity maps has an influence on the prioritization of areas for conservation. Nevertheless, parameterization and optimization of resistance surfaces in a biologically and ecologically relevant way for functional connectivity across species remains a nontrivial challenge41, and a better understanding of the relative importance of these factors is needed. Given a resistance surface, connectivity can be evaluated either by least cost path analysis24 or by quantifying current density based on circuit theory32,42. The latter approach is commonly used in landscape ecology and genetics studies32,42 as it allows for multiple movement pathways and varying degrees of corridor use across the landscape43. This makes it possible to investigate multiple corridor routing options44.
The spatial resolution of resistance maps has been shown to affect the accuracy of resulting connectivity maps through its effect on many landscape pattern metrics (e.g.45,46,47), including connectivity metrics48. Indeed, increasing spatial resolution (grain size) has a large effect on the accuracy of circuit-based connectivity estimates37. However, until recently49, computational limitations have precluded the use of circuit theory models to compute connectivity for large-extent, high-resolution maps29,50. Consequently, wall-to-wall, large-scale, and fine-grain computation of connectivity maps using circuit theory remains largely understudied, despite of increasing demand and recent expansions of this modelling approach10,28,34,51. Recent computational advances, namely the availability of GFlow49 and the new implementation of Circuitscape52 in Julia, have overcome previous computational limitations of Circuitscape and provide an important opportunity for developing and testing large-scale connectivity models based on current density.
Here we (i) propose a conceptual framework for species-agnostic connectivity modelling based on current density; (ii) discuss conceptual and computational decisions involved in implementing the approach; and (iii) assess the degree of uncertainty related to these decisions. The applied goal is to derive one main wall-to-wall map of current density as a measure of species-agnostic connectivity for Alberta based on the degree of human modification. The province of Alberta, Canada, is aiming to integrate an index of landscape connectivity into their biodiversity and ecosystem services assessment framework53. The Alberta Biodiversity Monitoring Institute (ABMI) is systematically collecting data on both human footprints and biodiversity for the province of Alberta at a level of detail, spatial extent, and spatiotemporal resolution that are unique at least for North America54 (see also the ABMI’s 10-year Science and Program Review, https://abmi10years.ca). Alberta, with a total area of 661,848 km2, has a human population of over 4 million (2016 census) and one of the biggest economies in Canada, including an expanding petroleum industry and significant contributions from agriculture, forestry, and tourism55. Because of rapid growth in population and economic activity, Alberta’s landscapes have changed noticeably over recent decades, and the rate of development is expected to increase further56, adding unprecedented pressure on remaining natural areas and low-intensity land uses57,58. Alberta defined a Land-use Framework53,59 to manage and sustain a growing economy, while balancing this with Albertan’s social and environmental goals. To improve land-use planning and biodiversity management in Alberta, policy makers, landowners, and other stakeholders are thus in need of new tools to evaluate the ecological value of ecosystems and landscapes, including an assessment of the contribution of an area to larger-scale connectivity, so as to compare the impact of alternative development scenarios and prioritize areas for conservation and restoration.
After deriving a main wall-to-wall map of current density as a measure of species-agnostic connectivity for Alberta based on the degree of human modification, we use this example to compare the contributions of different factors to uncertainty. We thus assess the sensitivity of this map to (i) the definition of the degree of human modification (i.e., the relative weight given to degree of physical human footprint vs. intensity of human use), (ii) the decision whether water bodies are considered barriers to movement, and (iii) the scaling function used to translate degree of human modification into resistance values. While the influence of most of these parameters on connectivity patterns has been previously studied independently, we aim to directly compare their contributions to uncertainty. We further (iv) evaluate the effect of such uncertainty on the classification of cells as being important or less important for maintaining larger-scale connectivity using different cut-off values. This study prepares the ground for future research to evaluate model validity with Alberta’s extensive biodiversity monitoring data.
Conceptual framework
We argue that a species-agnostic approach is conceptually better suited for integrating landscape connectivity into land-use planning, whereas a focal-species approach is better suited for conservation management (Fig. 1). Understanding the difference between these perspectives can help clarify conceptual differences, guide researchers in making decisions about how to model connectivity, and inform practitioners about the potential and limitations of resulting maps. Conservation is often based on focal species, e.g., in species-at-risk management, where it is paramount to adopt an organism perspective60 to define critical habitat and to consider the organism’s ability to move between habitat patches. This will result in a model of potential functional connectivity for the specific organism of interest, and a multi-species model can be derived by overlaying models for a representative suite of species10,22. In contrast, land-use planning focuses on the sustainable development of multi-functional landscapes. In this human-centred perspective, land parcel ownership and administrative boundaries define the relevant spatial scale and the degree to which landscape development can be influenced by policy, which in the case of Alberta includes the introduction of ecosystem services and biodiversity markets that play a major role in balancing environmental considerations with socio-economic drivers59. Regarding landscape connectivity, the focus thus lies on how human landscape alteration affects the connectivity of the remaining natural heritage system and how to compare the expected effect of local development alternatives on larger-scale connectivity. This focus is highly compatible with the modelling of connectivity based on human modification, which will result in a model of structural landscape connectivity.
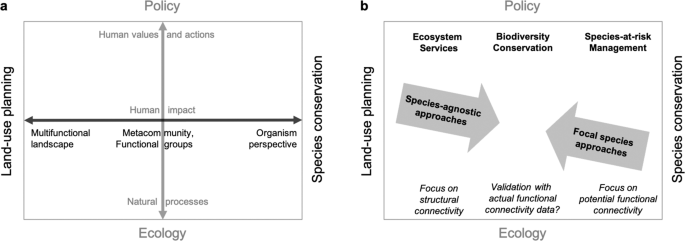
Conceptual framework: different perspectives on the landscape (a) and corresponding goals and approaches to connectivity modelling (b). The horizontal axis (black) illustrates how the landscape definition shifts from a multi-functional landscape in the context of land-use planning to an organism perspective in the context of species conservation. The vertical axis (grey) illustrates how the focus shifts from natural processes in basic ecology to human values and actions in a policy context. These values are reflected in policy that aims to preserve ecosystem services, biodiversity in general, or designated species of concern. Species-agnostic modelling of landscape connectivity may be most suitable in a land-use planning context, whereas focal species approaches may be most suitable in a conservation management context. When biodiversity data are available for many taxa, multi-species overlay can also be an efficient option to model connectivity for land-use planning and conservation programs for a representative suite of species. Validation of structural connectivity or potential functional connectivity based models could also be done with appropriate biological data, to assess actual functional connectivity.
A coarse-filter approach to biodiversity conservation management falls somewhere in-between61,62. The focus in this approach lies on preserving a community, such as grassland, wetland, or forest interior species, by maintaining a sufficient amount, quality, and connectivity of the respective ecosystem in the landscape. This implies that the species within a community have similar needs and characteristics, including their ability to move between habitat patches. Regarding landscape connectivity, we suggest that species-level data, such as mark-recapture, radio-telemetry, and molecular genetics data, which provide a direct measure of actual functional connectivity, may be used to test and compare species-agnostic (top-down) or multi-species (bottom-up) connectivity models. At the community-level, however, data such as biodiversity monitoring that is systematically collected across a large geographic area can be used as indirect measures of functional connectivity. If a connectivity model can explain, e.g., variation in species composition that remains unexplained by local site characteristics, this may indicate that it successfully captured the shared response of a broad range of species to human landscape modification. Note that there is a lack of empirical studies that compare a general-use connectivity assessment based on the overlay of many species-specific models across taxonomic groups to a species-agnostic model.
Another distinction occurs along the vertical axis in Fig. 1a: while ecology primarily considers natural processes, policy addresses human values and actions. The two perspectives meet in the consideration of the human impact on ecosystems. Based on this framework, a species-agnostic approach to modelling landscape connectivity quantifies how human actions and their manifestation as non-natural landscape features constrain ecological flow across the underlying natural fabric of the landscape. The main goals of this approach are to identify critical linkages and to support the evaluation of alternative development scenarios, with the general aim of maintaining ecosystem services and biodiversity in the context of sustainable development. This cannot replace fine-filter approaches for specific conservation goals, such as species-at-risk management. Note also that ecosystems can have an impact on humans as well, such as the spread of animal-transmitted diseases (e.g., West-Nile virus or Lyme disease). Here, we focus on human impacts on natural ecosystems.
To implement a species-agnostic approach, resistance values are assigned to non-natural landscape features based on expert opinion33. Conceptually, this involves two steps: (1) quantifying the degree of human modification, and (2) using a scaling function to assign resistance values based on the degree of human modification. We argue that beyond an inevitable element of subjectivity in expert-defined values, different interpretations of what constitutes degree of human modification can result in considerable differences between values. For instance, two types of roads may have the same physical footprint in terms of deviation from natural conditions (impermeable surface) but differ vastly in their intensity of use (traffic volume), which will affect e.g. road mortality rates. The relative scaling of resistance values is known to have a large effect on connectivity modelling, and a sharp contrast between low, medium or high resistance values has previously been suggested and used34,38,39.
In addition to human modification, some authors including Dickson et al.34 also considered natural barriers to the movement of terrestrial organisms and applied resistance values to features like water bodies and topography. Conceptually, this introduced an element of potential functional connectivity as it makes assumptions about the movement ability of organisms. In practice, this raises the concern that, e.g., water bodies may be traversable for some terrestrial species but not for others. Specifically, water bodies are a special case in such a framework as they are not a terrestrial feature, and while they are mostly natural, some authors have treated them as a barrier. However, in the context of species-agnostic models, much less thought has been given to the decision how to conceptualize water bodies than to other factors.
Here, we define a conceptually justified range of variation for the three factors identified above, with the goal of directly comparing their relative importance, as they have been addressed independently in previous studies on species-agnostic connectivity modelling: (i) the assignment of degree of human modification values by degree of deviation from natural conditions (footprint) or by the intensity of use; (ii) the assignment of resistance value to water bodies between minimum (natural) and a value close to the maximum of all human features combined (near-complete barrier); and (iii) the choice of scaling function resulting in a weak or strong contrast between non-natural and natural landscape features. We use intermediate values33 to create the main map, and we assess uncertainty in a factorial design using the extremes of each factor.
Circuit theory models connectivity as current density, where areas with high current density are interpreted as contributing to larger-scale landscape connectivity and thus ‘ecological flow’ across the study area32,42. We thus assess the contribution of each factor to variation among maps in the quantification of current density as the contribution of a cell to large-scale connectivity. We argue that visual interpretation of current density maps is affected by the choice of colour ramp, which should reflect a conscious decision about how cells with important contribution to large-scale connectivity (high current density values) are identified. J. Bowman (Trent University, pers. comm.) suggested that visualizing and interpreting current density values that have values higher than one standard deviation above the map mean (i.e., z scores> 1) as contributing areas provides a good balance for application. Here, we further vary this cut-off between the mean (i.e., z scores> 0) and two standard deviations above the mean (i.e., z scores> 2) to assess whether the uncertainty related to the factors above varies with the cut-off level used.
Source: Ecology - nature.com

