Database and conventional phylogenetic analysis
A 628 bp-long sequence of the control region of mtDNA were sequenced from 193 Moroccan individuals, belonging to 6 breeds (access numbers in GenBank: MN229085-MN229277). To enrich the database, the use of these sequences as seeds allowed retrieving from NCBI Blastn tool 652 foreign sequences, distributed in 9 Iberian, 7 Italian, and 10 Oriental breeds. There are two non-native breeds: the Lacaune of Italy and the German Merino. The number of individuals per breed, their origin and their access number are indicated in Suppl. S1. 1.
The phylogenetic analysis generated a tree encompassing a total of 845 individuals, rooted by individuals of C and E haplogroups according to Meadows et al.20, and displayed in Suppl. S2. Among the 193 Moroccan individuals, 185 were found to belong to haplogroup B, i.e. 96% (Suppl S1. 2). It is worth noting that at the base of the C and E haplogroup clade, there is a series of four embedded Moroccan individuals (GenBank access numbers: MN229117, MN229266, MN229115, MN229259). Similarly, there are two sequences of Moroccan breeds also at the base of the A haplogroup clade (Fig. 1) (GenBank access numbers: MN229219, MN229173). Conversely, at the root of the B haplogroup clade, there are three Moroccan individuals associated to Portuguese Churra Badana breed, two others to Italian Merinizzata Italiana breed, and the last one to the Turkish Herik breed.
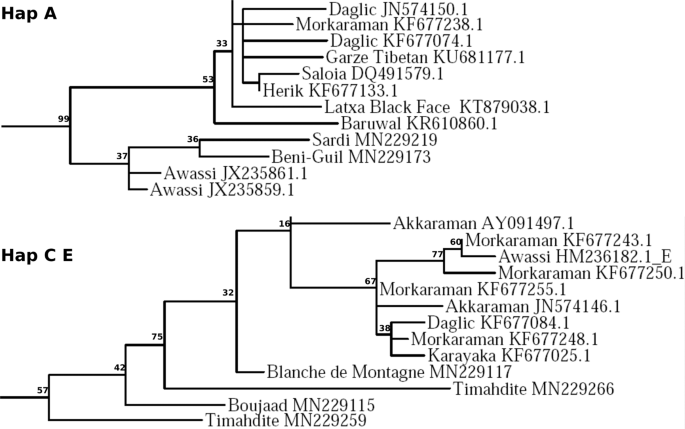
Position of the Moroccan breeds at the base of haplogroups A and C E. The bootstrap supports were calculated from 500 iterations.
Variation in the composition and connectivity among regions and breeds
The treatment of the topology information at the level 1, i.e. terminal branches, using Newick-Extra produced the table of affinities between breeds (Suppl. S1. 3), and then their proportions for each breed. As a first approach, the non-metric multidimensional scaling (N-MDS) analysis (Fig. 2a) revealed the isolated position of Oriental breeds, and the connections between Moroccan and Iberian breeds on one hand, and between Iberian and Italian breeds on the other hand. A more detailed view was given by the Cluster analysis (Fig. 2b), which highlighted three groups: (i) Oriental breeds, (ii) Moroccan breeds associated to four Portuguese breeds, and (iii) a mix of Iberian and Italian breeds.
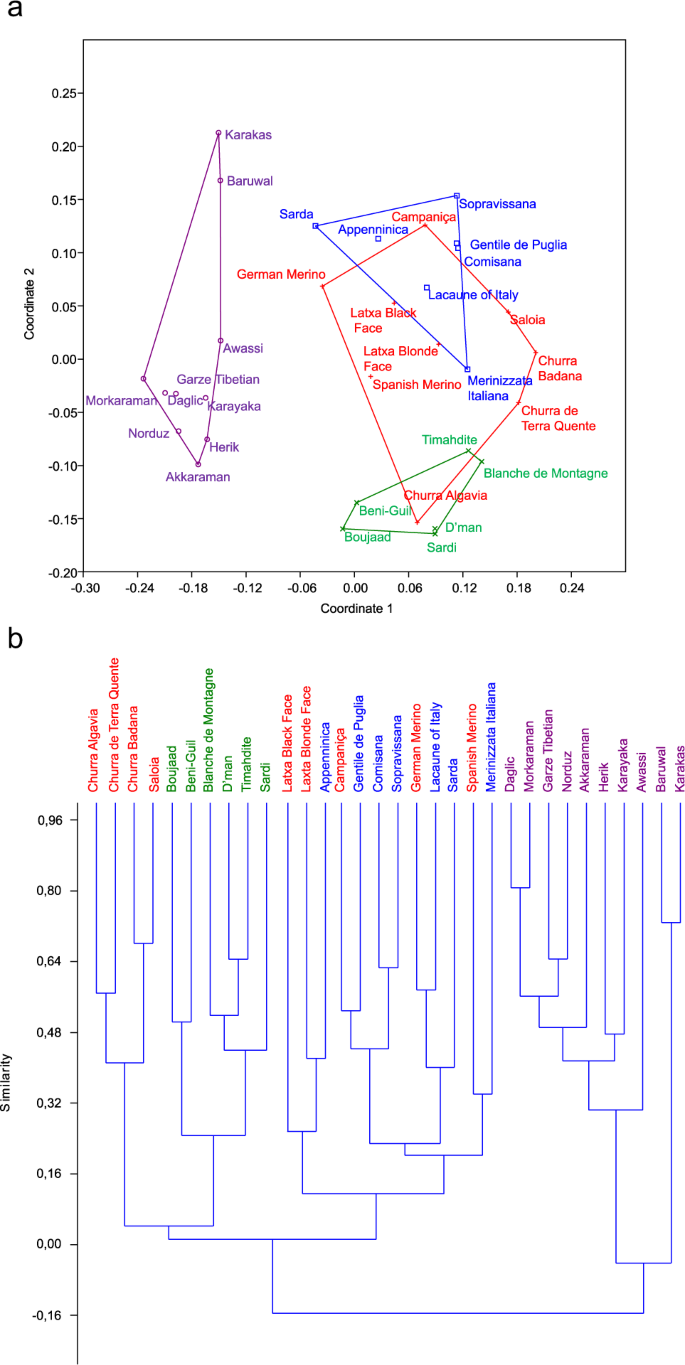
Relationship between breeds according to their geographic origin, from Newick-Extra program. (a) N-MDS analysis showing the intermediate position of Iberian breeds among western Mediterranean ones. (b) Cluster analysis highlighting the proximity between Moroccan and Portuguese breeds. Violet = Oriental breeds, Blue = Italian breeds, Red = Iberian breeds, Green = Moroccan breeds. The figure was drawn using PAST software v. 2.97, available at https://folk.uio.no/ohammer/past/.
In the same perspective of studying connectivity and kinship between populations, the genetic distances Dxy and Da between breeds (Suppl. S1. 4) were visualized by NMDS. In both graphs (Suppl. S3. A-B), the Moroccan, Iberian and Italian breed ellipses were strongly superposed, while the Oriental breeds were scattered for Dxy genetic distance and relatively grouped for the Da genetic distance. These results were confirmed by Cluster Analyses (Suppl. S3. C-D).
The use of Network software v.5.0.0.1 showed seven groups. On the left and the right sides, two groups gather individuals of haplogroups C-E and A respectively, dominated by animals of Oriental breeds (Fig. 3). The situation concerning haplogroup B is more complex as the individuals were organized in four groups: one Italian, one Iberian and two Moroccan ones, named 1 and 2, represented by 119 and 32 individuals respectively, making 62% and 17% of the actual Moroccan genetic background. The Iberian one was localized in the center and closer to the Moroccan group 1 than to the Italian one. The Moroccan group 2 was linked to the Italian one and the ensemble composed of individuals of haplogroups C and E. At a closer look, it appears that (i) the Italian and Iberian groups included 20 and 14 Moroccan individuals respectively, corresponding to c. 10% and 7% each of the total from Morocco; (ii) moreover, the Italian group also contained several Iberian (Spanish Merinos only) and Oriental individuals); (iii) the Iberian group itself hosted individuals of Oriental, Italian, and of the Moroccan group 1. As regard haplogroup D, composed of a few oriental individuals, its connections concern haplogroup A group and two ensembles belonging to haplogroup B, i.e. Iberian group and Moroccan group 1.
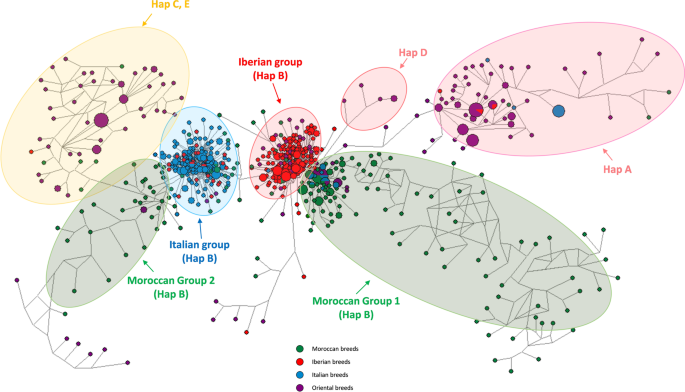
The individuals are organized in four haplogroup ensembles. The haplogroup B is dominant and comprises the Italian, the Iberian and the two Moroccan groups. The figure was drawn using Network software v.5.0.0.1, available on the website: http://www.fluxus-engineering.com/.
To test whether the genetic composition of the three western Mediterranean areas (Moroccan, Iberian, and Italian) was different, the AMOVA revealed significant molecular differences among breeds and within breeds for the 3 groups. Regardless of the compared groups, the main source of variation lay within populations, exceeding 96%. More interestingly, it appeared that there were significant differences between all the combinations of groups, except between Iberian and Italian ones. At a closer look, the difference between Moroccan and Iberian breeds was lower than between Moroccan and Italian ones (Suppl. S4. A).
In summary, the structuration of the western Mediterranean populations was relatively concordant with the results of procedures involving the Network, the AMOVA and Newick-Extra program, while the Da and Dxy genetic distances provided a confusing view. To test specifically the level of compatibility between the Newick-Extra program and the Network methods, we generated 2 matrices that presented the percentages of connections between each Moroccan breed and the other breeds according to their geographical origin (Moroccan, Iberian, Italian or Oriental). In contrary to the Newick program for which this counting was straightforward, the count of connections from the network needed to be done manually. For each Moroccan breed, the numbers of individuals belonging to Italian, Iberian, Moroccan (1, 2 and 3) or Oriental (Haplogroups A, C, D, and E) were summed (Suppl. S1. 5). The correlation between the two matrices was highly significant (N = 24, r = 0.93, p < 0.001).
For each Moroccan breed, the percentages of affinity (Fig. 4) showed contrasted situations toward Italian, Iberian and Oriental breeds. Beni-Guil breed had the highest proportion of influence from foreign countries (about a third), especially from Iberia, while D’man breed affinity was specifically Moroccan. The remaining Moroccan breeds had about a fifth relationship with foreign breeds, the Timahdite being the only breed devoid of connection with oriental breeds. There were moderate connections between Moroccan breeds themselves, except for the D’man, which showed more than 20% of relationship with the Timahdite, the Blanche de Montagne and the Sardi breeds. The analysis of the phylogenetic tree (Suppl. S2) indicated that all these connections were situated in the terminal branches of haplogroup B clade, and thus corresponded to recent exchange events.
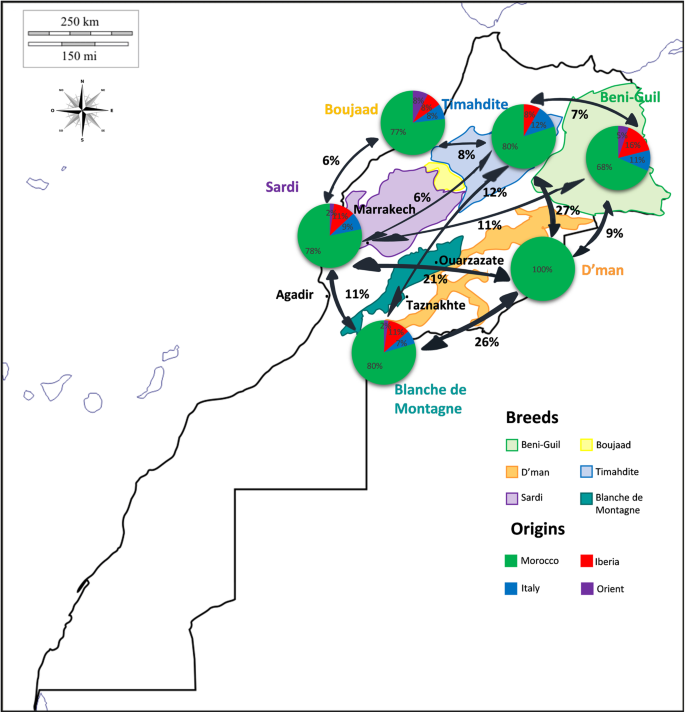
Map of connections between each Moroccan breed and (i) other Moroccan ones, and (ii) breeds of other Mediterranean areas.
Climatic parameters
As it seems that the Italian, Iberian and Moroccan regions are centers of differentiation of sheep populations, it is useful to characterize the climatic constraints that exist in these three regions. The ANOVA conducted on the sum of localities recorded for each breed within each region lead to significant differences for most parameters (Suppl. S1. 6 and S3. E). As the altitudes recorded in the breeding areas in Morocco are higher than in Italy and Iberia, the winter temperatures expressed by m are colder. Given the south position of Morocco, the summer temperatures expressed by M are higher, leading to a decrease in rainfall (R) and vegetal productivity (Q3).
With regard to the breeds themselves, the mean values for climatic parameters were gathered in S1. 7.
Genetic variability parameters
Among the groups defined from haplogroup B, it appeared that the Moroccan group 1 had a significant lower haplotype diversity Hd than the Iberian and Italian ones, but a higher nucleotide diversity π in relation to numerous singletons. At the level of breeds, it was expected that the variations in genetic diversity between Moroccan breeds followed the degree of affinity to foreign ones showed in Fig. 4. Effectively, the haplotype diversity Hd had its highest and lowest values in Beni-Guil and D’man, respectively (Suppl. S4. B., Suppl S1. 2 for all breeds).
Neutrality tests and climatic constraints
The calculated neutrality tests Fu and Li F* and D* among the four ensembles defined from haplogroup B, indicate lower significant values in Moroccan group 2 and Italian group than in the two other ones. These same tests applied to Moroccan breeds highlighted the Boujaad, the Blanche de Montagne and the Beni-Guil breeds, partially retrieved using Tajima D test. In contrast, Fu’s Fs revealed that all the Moroccan breeds were significantly out of neutrality. As it is difficult to quantify a direct influence of human selection by neutrality tests, it seems easier to investigate the effects of environmental constraints, especially the climatic parameters. For this purpose, a synthetic database was constructed by including both climatic and genetic data (Suppl. S1. 7). The relationships among all the climatic and genetic variables were well represented on the PCA first plan (70.02% of variance) (Fig. 5). The pluviometric coefficient Q3 and annual rainfall R varied in opposite to the temperature variables (M, T max, M – m) and altitude. In contrast, the variables m and T min seems to vary in the same direction as most genetic variables except Hd. With regard to breeds, the Moroccan ones and the Iberian Spanish Merino and Churra Badana breeds were located towards high temperature and low rainfall variables. Another set of breeds was clearly associated to high rainfall, e.g. Sarda, Sopravissana, Saloia, Latxa Blonde and Black face. There is another set of three breeds (Campaniça, Churra Algarvia and Comisana) sharing high values of neutrality test variables. The result of correlation indicated that the variables T min and m were positively and significantly associated to π, Fu and Li F* and D*, and to some extent to D of Tajima (Suppl. S1. 7, Suppl. S3. F–H). If the breeds for which the significant neutrality tests Fu and Li F* and D* were considered (Blanche de Montagne, Boujaad and Beni-Guil), it appears that they were located in the PCA plan towards higher altitudes, low minimal temperatures (m and T min) and higher maximal temperatures (M and T max).
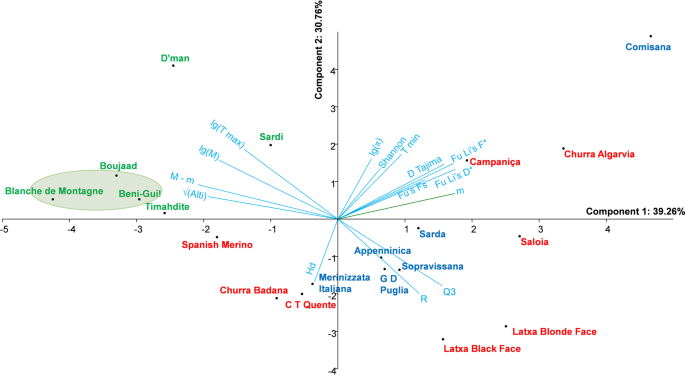
PCA of the climatic and genetic parameters. The ellipse gathers the breed significantly under selection (Fu and Li D* and F* parameters). Color code: see Fig. 2A. The figure was drawn using PAST software v. 2.97, available at https://folk.uio.no/ohammer/past/.
Expansion time
To assess the prospective expansion events of western Mediterranean native breeds, the calculation of mismatch distribution revealed that the four groups of haplogroup B and breeds followed the coalescent simulation (p-value of SSD test >0.1, Suppl. S3. I,J), except the limit case of Comisana (p-value of SSD test = 0.08). The τ variations did not correlate to any climatic parameters (Suppl. S1. 7). From the first method of expansion dating, the yearly mutation rate could be estimated at 3.4 × 10−5 and 2.3 × 10−5 using 1.26 and 1.72 Myrs ancestors, respectively, resulting in expansion times at 140,000 and 207,000 years for the D’man breed for example. From the second method using archaeological records of Italy and Iberia, it appears that the Moroccan group 1 seems to have expanded later than Iberian one around 7,100 years B.P., whereas the Moroccan 2 group showed an earlier expansion than the Italian one itself around 8,600 years B.P. Due to the low population size of Moroccan individuals recorded in Italian and Iberian groups, no mismatch analysis could be done. With regard to the Moroccan breeds, the northern ones (Timahdite, Sardi, Beni-Guil and Boujaad) are in the range of c. 6,700–7,400 years B.P., while the southern ones are associated to 7,700 years B.P. for the Blanche de Montagne and 8,400 years B.P. for the D’man (Suppl. S1. 8).
Source: Ecology - nature.com

