Hypolithic horizon: types and transitions
Hypolithic horizons most widely occurred in soils of the valley floors and flanks, which were elevated at least ten centimeters above the zone of meltwater streams at the peak of the warm season. We observed two distinct morphotypes of hypolithic horizons (Fig. 2): (1) cyanobacteria-dominated, which comprised biofilm-covered sandy particles several centimeters beneath the gravel pavement and patchy biofilms on the undersides of pavement components and (2) moss-dominated, which comprised living moss biomass beneath the pavement and peat material below it (rarely thicker than 5 cm). We also observed transitional series of hypolithic horizon, that were represented by both cyanobacteria- and moss-dominated morphotypes with the latter one proliferating into epiedaphic environment between unevenly distributed pavement components. We did not find a separate fungus-dominated morphotype like in studies of the Dry Valleys soils20,35, although fungi mycelium and spores were previously reported from hypolithic habitats of the Larsemann Hills46. We also recognized dark colored horizons with filamentous biofilms on sandy particles that were buried in the microtopography traps immediately below contemporary hypolithic horizons dominated by cyanobacteria (Fig. 3).

Major types of hypolithic organogenous horizons produced by BSC in soils at the valley floors in the Larsemann Hills: (a) patterned ground controls moisture distribution and transitions from cryptic hypolithic organogenous horizons (on polygons) to patchy epiedaphic organogenous horizons (in troughs); (b) plane and soil profile view on various types of hypolithic horizons on polygons; (c) soil profile in the transitional zone between hypolithic horizon produced by cyanobacteria- and moss-dominated BSC (polygon periphery) to the patchy epiedaphic horizon produced by moss-dominated BSC (trough). The tape-measure on photos is in cm.
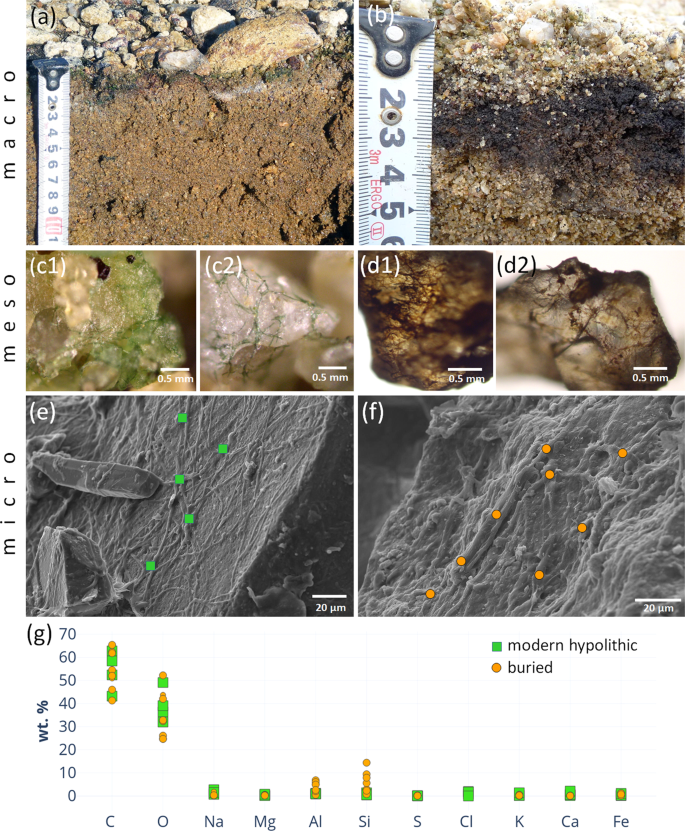
Morphology and composition of cyanobacteria-dominated hypolithic horizons at various scales: (a) cyanobacteria-dominated morphotype of the high center polygon at the wet valley floor; (b) dark colored sandy material buried under contemporary cyanobacteria-dominated hypolithic horizon in microtopography trap at the polygon periphery; (c1, c2) filamentous morphology of cyanobacterial biofilm from the contemporary hypolithic horizon, optical microscopy, reflected light; (d1, d2) − filamentous morphology of presumably cyanobacterial biofilm from the buried horizon found in microtopography trap at the polygon periphery, optical microscopy, reflected light; (e) SEM-SE image of the contemporary cyanobacterial biofilm on the surface of feldspar particle; (f) SEM-SE image of the presumably cyanobacterial biofilm on the surface of feldspar particle, buried horizon found in microtopography trap at the polygon periphery; (g) elemental composition of modern hypolithic and buried horizon as suggested by the EDX data, orange circles and green squares indicate values obtained from individual EDX measurements, corresponding acquisition sites are marked at SEM-SE images e, f.
Occurrence and transitions between the two morphotypes of hypolithic horizons depended on the topography gradients, which redistributed meltwater when available and enabled development of various BSC types. We observed lateral hypolithic patterns both on the scale of tens of centimeters due to cryogenic microtopography (modified by rill erosion near the thalweg zone) and on the scale of several meters due to the mesotopography gradient from the valley floor to its gentle slopes. At the high flanks of the valley, the macroscopic organogenous horizons (even the cryptic ones) were almost absent giving a way to the barren ahumic soils with no visible traces of photoautotrophs. At the valleys floors, the patterned ground often controlled moisture distribution and transitions from cryptic hypolithic organogenous horizons on polygons to patchy epiedaphic organogenous horizons in troughs (Fig. 2a). Polygon periphery was a transitional zone between hypolithic horizon produced by cyanobacteria- and moss-dominated BSC to the patchy epiedaphic horizon produced by moss-dominated BSC in trough (Fig. 2b,c).
We determined a lateral gradient in the gravimetric moisture content in the top 5 cm of soil at the 4 × 11 m moisture mapping site in the valley 1. This gradient arose from the 2 m difference in elevation between the valley floor and its east-southeast flank (Fig. S5c) that caused meltwater redistribution. We found that observed BSC types occurred within different moisture ranges in 4 zones (Fig. 4). There were no macroscopic biofilms found on the undersides of pavement components and in its sandy bedding in a range of 0.4–3.7 wt. % of moisture (zone 1). Hypolithic cyanobacteria-dominated morphotype was present when moisture content varied between 3.8–7.8 wt. % (zone 2). The first moss-dominated hypolithic horizons scarcely appeared when soil moisture content approached 7.0 wt. %, then became continuous and mixed with epiedaphic moss-dominated horizons with up to 27.1 wt. % of soil moisture (zone 3). Tongues of amphibian cyanobacteria- and algae-dominated BSC appeared in a range of 18.7–46.1 wt. % (zone 4). The cryogenic microtopography (slightly modified by rill erosion) in zone 2 and partially in zone 3 caused the interplay of the two moisture ranges: 3.8–7.8 wt. % that was correspondent to the cyanobacteria-dominated hypolithic horizons on polygons and 7.0–17.0 wt. % that was common to the transitional series and moss-dominated horizons in troughs. It is well established that hypolithic colonization is highly correlated with water availability at the global scale63. The gradients in moisture content estimated in this case study from the Larsemann Hills also gave a very good explanation to the observed BSC sequences and associated carbon-rich horizons both on the micro- and mesotopography levels. At the same time, we are aware that caution is needed in attributing the field measured moisture content directly to the physiological processes in BSC and the soil organic matter build-up since it can vary a lot in time. Previously Ellis-Evans48 found subsurface communities were still present in lithosols of the Larsemann Hills within 1–2 wt. % of soil moisture and the laboratory experiments with cyanobacterial hypoliths of hot deserts showed that primary production required a minimum of 15 wt. % of soil moisture31.
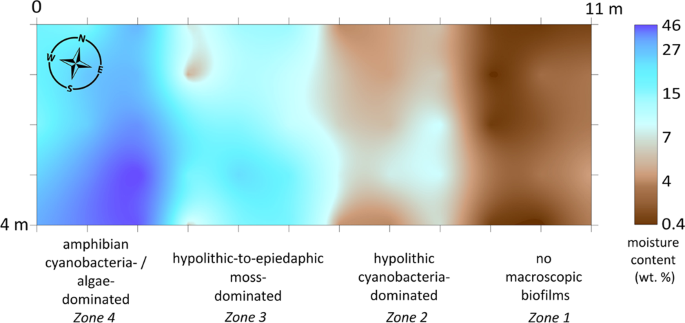
Spatial distribution of gravimetric moisture content (top 5 cm of soil) and different types of BSC at the 4 × 11 m moisture mapping site (60 measuring points) in the valley 1 of the Larsemann Hills (69.39004°S, 76.40427°E; for location in the valley see also Fig. 1). Note: (1) when the gravel pavement was present, the samples were collected directly beneath it in the sandy bedding; (2) in case of epiedaphic moss-dominated horizons the moisture content was measured in peat and mineral fine earth immediately beneath the living biomass.
According to the field observations in 2010 and 2016, the patterned ground at the wet valley floors had a quasistationary surface. Along with cryogenic translocation of material due to the formation and decay of polygons, there were clear signs of the lateral transport of fine earth at the microtopography scale from the high centers to the more wind-sheltered troughs. We hypothesize that these processes enabled deeper burial of organogenous material, which was originally produced in hypolithic/epiedaphic environment, and therefore led to the enrichment of soil in organic matter. We observed the possible long-term consequence of these processes as the formation of dark carbon-enriched layers few centimeters beneath contemporary hypolithic horizons (Fig. 3b). These buried layers consisted of the sand particles enclosed in biofilms. The biofilms had a distinct filamentous morphology (Fig. 3d,f), which was very similar to the morphology of cyanobacterial biofilms (Fig. 3c,e) from the hypolithic horizon above. The elemental composition obtained from EDX spectra was also in the agreement between contemporary and buried layers (Fig. 3g), although Al and Si abundances were higher in the buried material. The buried biofilms were thick (10–15 µm) and the biogenic filaments were embedded in the amorphous material. The thickness of biogenic coating on the sandy particle of more than 10 µm secured that the Al-Si signal extracted by SEM-EDX originated from the coating itself, but not from the mineral background. This indicates biofilm fossilization and possible close integration of organic matter and amorphous Al/Si-containing species that made this biofilm-to-mineral association more stable.
Organic matter in soils with hypolithic horizons
A set of 180 samples was processed to obtain the total organic carbon (TOC) and the total nitrogen (TN) concentrations, as well as TOC/TN ratios in soils with hypolithic horizons located at the wet valley floors in the Larsemann Hills. The whole data array revealed that TOC concentrations were never <0.04% with a mean of 0.68 ± 0.85% (±1 standard deviation) and TN concentrations ranged from 0 to 0.38% with a mean of 0.05 ± 0.07%, indicating that values were widely spread out. The lowest values were recorded in barren ahumic soils with no macroscopic traces of BSC (even in the cryptic niches), which contained 0.04–0.07% TOC and 0.01–0.02% TN or no detected nitrogen at all (Table S1). At a detailed scale there were distinct differences in concentrations of carbon, nitrogen and their ratios within hypolithic horizons themselves, and also between hypolithic, buried organogenous and subsoil horizons. Moss-dominated horizons expectedly had higher TOC and TN concentrations than cyanobacteria-dominated ones (Fig. 5a,b; Table S2): 2.75 ± 1.10% vs. 0.44 ± 0.31% for TOC and 0.22 ± 0.09% vs. 0.04 ± 0.03% for TN, respectively. The mean of TOC/TN ratio for the whole data array (excluding the cases when nitrogen has not been detected) was 14.9 ± 7.1. The TOC/TN ratio in the moss-dominated horizons was rather low with a mean of 12.7 ± 1.2 and a narrow range of 10.8–14.7, which could be explained by the presence of the well-decomposed and homogenized organic matter, mainly represented by peat with hardly discernible or no plant structure. The TOC/TN values in the cyanobacteria-dominated horizons varied between 4.0 and 43.4 with a mean of 15.3 ± 7.5 and demonstrated a skewed distribution with some distant outliers from the bulk of the data (Fig. 5c). TOC/TN ratios can be misleading if the organic matter content is low and the stoichiometry patterns are controlled by unknown variables, e.g., the legacy carbon or mineral impurities. However, when the cyanobacteria-dominated BSC measurably contributes to the soil organic matter (up to 1.34% TOC in our dataset) the elevated TOC/TN values could indicate the extensive production of the C-rich extracellular polysaccharides64 common to the subaerial cyanobacterial biofilms65 especially in the extreme environments. The organic matter properties of the moss- and cyanobacteria-dominated horizons are well discriminated on the TOC–TOC/TN plot (Fig. 5e): the moss-dominated series are enriched in TOC while the TOC/TN range is consistent.
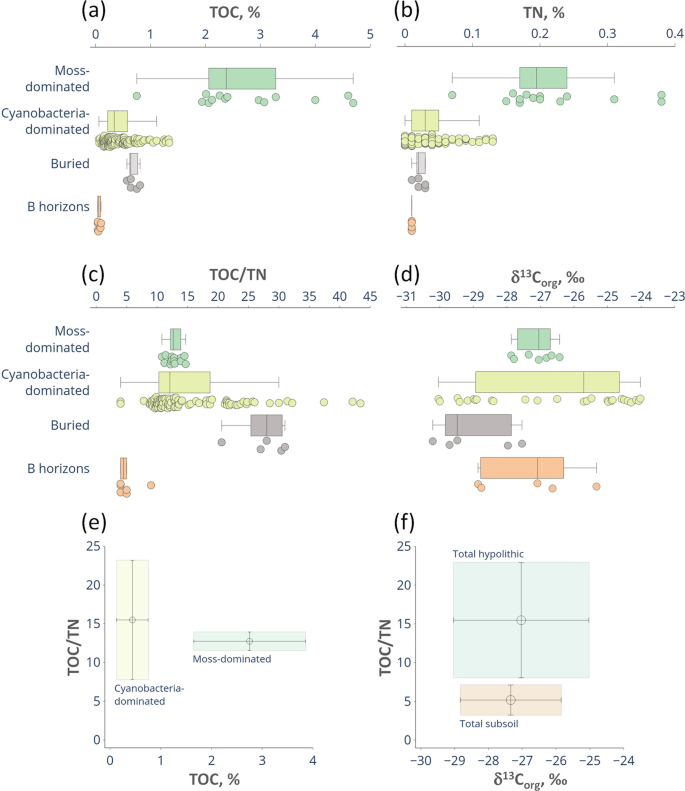
TOC (a) and TN (b) concentrations, TOC/TN (c), carbon stable isotope ratios δ13Corg (d), TOC-TOC/TN (e) and δ13Corg-TOC/TN (f) plots in various types of soil horizons at the valley floors, Larsemann Hills. Elements of the box plots represent minimum excluding outliers, first quartile, median, third quartile, maximum excluding outliers, while the points represent individual values of the whole data array including outliers. The values plotted at (e) and (f) are the means of δ13Corg and TOC/TN values and the bars are standard deviations.
Previously, the imbalanced stoichiometry (low C, high N) with a very little biotic control over bulk element ratios was suggested as a general property of soils in extreme environments of Victoria Land in Antarctica66. Our findings in the milder setting of the Larsemann Hills suggest a more pronounced biotic influence on TOC, TN and their ratio at least in the hypolithic horizons dominated by mosses and cyanobacteria.
The δ13Corg values (n = 44) ranged from −30.2 to −24.0‰ (Fig. 5d) with the mean of −27.1 ± 1.9‰ The δ13Corg interval (mean ± 1 standard deviation) of the subsoil horizons stayed within the total interval of the topsoil hypolithic horizons and highlighted the possible link between hypolithic environment and deeper layers, while the intervals of TOC/TN ratios were clearly discriminated between hypolithic and subsoil horizons (Fig. 5f). Observed δ13Corg ranges are consistent with the subaerial biogenic sources of organic matter7 and this relates not only to the topsoil but also to the subsoil. Our dataset is in line with the δ13Corg ranges of the endolith-derived organic matter, lichen and moss communities3,7,67,68. We found that isotopic signature of the cyanobacteria-dominated hypolithic series of the Larsemann Hills varied in the wider range (−30.0 – −24.0‰), than for the moss-dominated ones (−27.9 – −26.4‰). The observed δ13Corg range of hypolithic horizons dominated by cyanobacteria is most close to the one in endolithic systems3,7, which are also dominated by cyanobacteria, often in the lichenized form. The occurrence of free-living or lichenized fungal communities in cyanobacteria-dominated hypolithic horizons of the Larsemann Hills is plausible and could explain why these horizons are more 13C-depleted than surficial cyanobacterial and algal communities7,68. Discrimination during biosynthesis in fungi could result in the lower δ13C values since fungi contain more structurally complex biopolymers than cyanobacteria or algae7. This is also consistent with the presence of mycelium in hypolithic horizons of the Larsemann Hills46, which, however, does not form distinct fungus-dominated morphotype.
Although the contemporary biotic influence on the soil organic carbon and its stable isotopes ratio is well pronounced in the Larsemann Hills, we cannot exclude the input from other sources of organic carbon, both in the hypolithic topsoil and the subsoil, e.g., redistributed organic matter from the paleo (legacy carbon) and modern lake sediments7,69,70.
Spatial patterns of TOC and TN at a detailed scale
TOC concentrations at the 10 × 10 m hypolithic site with 121 sampling points ranged from 0.06% to 4.69% and TN values varied between 0.01% and 0.38% (Fig. 6). TOC and TN spatial patterns often correlated and the elements abundances were the lowest in the middle part of the high center polygons, where biofilms were primarily present on the undersides of the gravels and more rarely extended deeper into the sandy bedding. The loci with the highest TOC and TN concentrations were mainly found in the trough areas (Fig. 6g) and were associated with the moss-dominated horizons. TOC values in troughs averaged at 1.07 ± 1.03% and at the high centers – 0.37 ± 0.28%, while TN in troughs averaged at 0.11 ± 0.09% and at the high centers – 0.04 ± 0.03%. Medians of TOC and TN concentrations had statistically significant difference between polygons and troughs (Table S2). A possible explanation of this pattern is that troughs provide better moisture conditions, and also serve as wind shelters at the microtopography level enabling proliferation of hypolithic BSC into the epiedaphic environment or even individual development of the epiedaphic BSC. Analysis of soil temperatures within 20 cm from the surface of the patterned ground showed that the austral summer temperatures in the trough are also more favorable than in the high center of the polygon. The average January temperature at a depth of 20 cm reached + 4.9 °C in trough versus +4.4 °C on high center, while the annual sum of T > 0 °C was 261 °C versus 229 °C, and the annual sum of temperatures >5 °C was 133 °C versus 78 °C, respectively. Increased soil temperatures and longer warm periods in troughs originate from the larger pore space between frost sorted coarse materials in comparison to high centers. The proliferation of mosses into epiedaphic environment in troughs induced by its favorable physical conditions also enables positive feedback on soil temperature regime from better absorption of solar radiation by patches of the dark moss biomass (if patchy moss would develop into continuous epiedaphic cover it could cause the opposite effect due to thermal insulation).
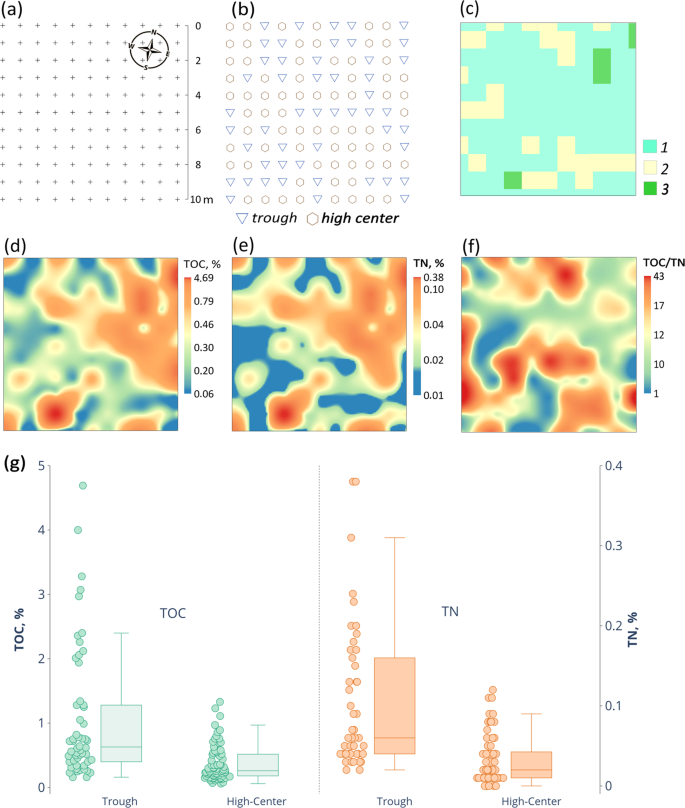
Spatial distribution of TOC, TN and their ratios at a detailed scale (Larsemann Hills, valley 1, 69.3901°S, 76.4039°E): (a) sampling design on the 10 × 10 m hypolithic site, crosses mark sampling points; (b) microtopography of the key site; (c) distribution of hypolithic horizons: 1 – cyanobacteria-dominated morphotype with hypolithic colonization of both the gravel undersides and sandy bedding of the gravel pavement; 2 – cyanobacteria-dominated morphotype with hypolithic colonization of only the gravel undersides; 3 – moss-dominated morphotype with mainly hypolithic colonization and patchy proliferation into epiedaphic environment; (d, e, f) spatial patterns of TOC, TN and TOC/TN, respectively; (g) TOC and TN concentrations in hypolithic horizons upon microtopography pattern. Elements of the box plots represent minimum excluding outliers, first quartile, median, third quartile, maximum excluding outliers, while the points represent individual values of the whole data array including outliers.
Overall, the detailed mapping at the 10 × 10 m hypolithic site has documented the presence in hypolithic environment of a continuous organogenous horizon that spreads for at least 100 m2 and even proliferates from the cryptic series to the open ones upon a patchy pattern. The most common was the hypolithic cyanobacteria-dominated morphotype which consisted of two subtypes: 1) with the colonization of the gravel undersides and 2) with the colonization of both the gravel undersides and the sandy bedding of the pavement. We suggest that troughs which display the highest TOC and TN concentrations are the hot spots of organic matter accumulation at a detailed scale upon a patterned ground template in the presence of meltwater. Potential mechanism of a longer carbon stabilization in troughs involves burial of organic matter by the fine earth eroded from the adjacent polygons or windblown from the more distant sources. This is one of the possible pathways for hypolith-derived organic matter to enrich the soil and produce the buried organogenous layers, e.g. those with dark filamentous biofilms as seen in (Figs. 3b,d1,d2,f).
Radiocarbon age of TOC
We summarize here the radiocarbon data on TOC from cyanobacteria-dominated hypolithic horizons, moss-dominated hypolithic and epiedaphic horizons, and also subsoil horizons (Table 1 and Fig. S3). Hypolithic and epiedaphic horizons contained a significant amount of modern carbon with the F14C(fraction modern)>1 in 4 out of 5 samples from the cyanobacteria-dominated series (max F14C = 1.168 ± 0.004), and in 2 out of 3 samples from the moss-dominated ones (max F14C = 1.121 ± 0.003). In the hypolithic horizons there were two samples with F14C < 1 corresponding to a conventional 14C age of 150 ± 20 yr BP (F14C = 0.982 ± 0.003) for TOC of cyanobacteria-dominated material and 1230 ± 45 yr BP (F14C = 0.858 ± 0.005) for TOC of the moss peat. The shallow B horizon with no macroscopic signs of biota revealed F14C > 1 at a depth of 5 cm, which suggests young carbon has leaked to the subsurface, e.g., during the meltwater percolation. The deeper B horizons were as old as 1840 ± 20 yr BP (10–15 cm) and 6690 ± 30 yr BP (20–25 cm) corresponding to F14C = 0.795 ± 0.002 and F14C = 0.435 ± 0.001. TOC in the shallow buried horizon with fossilized biofilms of presumably cyanobacterial origin (Figs. 3b,d1,d2,f) had 14C age of 1105 ± 25 yr BP (F14C = 0.872 ± 0.0025).
14C data indicates that organic carbon stored in hypolithic environments of the Larsemann Hills is relatively young. This is very different from the findings in the most extreme hyper-arid environments like Atacama Desert, where hypoliths establish long-living communities with ages of up to 12,000 years since the substrate colonization27. The water supply is crucial for high productivity of hypolithic communities and their age is positively correlated with aridity27,33. In the Larsemann Hills the hypolithic communities of the valley floors are relatively well supplied with meltwater in comparison to the driest locations in Atacama Desert or Dry Valleys. Previously Zazovskaya et al.55 also proposed the high dynamism of the soil surfaces due to strong winds and occasional erosion by meltwater as a possible explanation of young 14C ages of TOC in the topsoil, which one could assume should experience very slow C turnover times in the extreme environments of East Antarctica. Our radiocarbon dataset supports the hypothesis of several time-divergent carbon pools in soils of the Larsemann Hills with rather rapidly cycling carbon decoupled from the ancient one like it was previously proposed for the Dry Valleys soils4,71. While the topsoil in the Larsemann Hills has an overwhelming portion of modern TOC there are still 20–30% of cases where organic carbon is stabilized for ~100 years in cyanobacteria-dominated hypolithic horizons and ~1000 years in the moss peat of the epiedaphic/hypolithic organogenous horizons. We report here that the burial under a few centimeters of fine earth is an efficient mechanism to stabilize cyanobacterial biofilms for ages up to ~1000 years. 14C age of TOC in deeper horizons of soils points toward at least partial contribution of the legacy carbon.
CO2 efflux from soils
Carbon dioxide emission from the surface of soils (valley 1, Fig. 7) with the cyanobacteria-dominated hypolithic horizons was as low as 8.0 ± 0.7 mg C-CO2 m−2 hour−1 (± standard error of the mean). The appearance of mosses in the hypolithic environment with scattered proliferation on the day surface slightly shifted emission values to 12.4 ± 1.2 mg C-CO2 m−2 hour−1. This is obviously explained by the higher biomass of mosses, their enhanced moisture capacity and better warm-up of the dark epiedaphic patterns, all together stimulating heterotrophic activity. To better understand CO2 fluxes from soils with hypolithic horizons and emplace them on the landscape level we simultaneously measured the surface emission from the most carbon-rich and carbon-poor types of soils in valleys of the Larsemann Hills. We chose soils in the wind-sheltered depressions on the rocky slopes (see Fig. 1 for location) as the most advanced series with the 10 cm thick epiedaphic moss horizons. As the most carbon-poor background control, we employed the barren ahumic soils in the valley 1 with no macroscopic signs of any BSC. The soils in wind shelters had four times higher CO2 emission than the moss-dominated mixed hypolithic/epiedaphic series at the valley floors. At the same time, soils with cyanobacteria-dominated hypolithic horizons had less than 5 mg C-CO2 m−2 hour−1 increment in flux in comparison to the dry soils of higher valley flanks, which demonstrated no macroscopic BSC at all. Since the measurements were conducted simultaneously and the temperature and moisture content were comparable, this difference could be indicative of CO2 added by cyanobacteria-dominated hypolithic horizons to the CO2 flux apart from its abiogenic component. Shanhun et al.72 showed that biological respiration in the Dry Valleys accounts only for 25% of the measured CO2 flux from soil. In the milder environments of the Larsemann Hills, the input from biological respiration could be greater. We cannot completely exclude heterotrophic activity even in the driest (moisture content ~3%) ahumic soils, which contained in our dataset at least 0.04% TOC (Table S1).
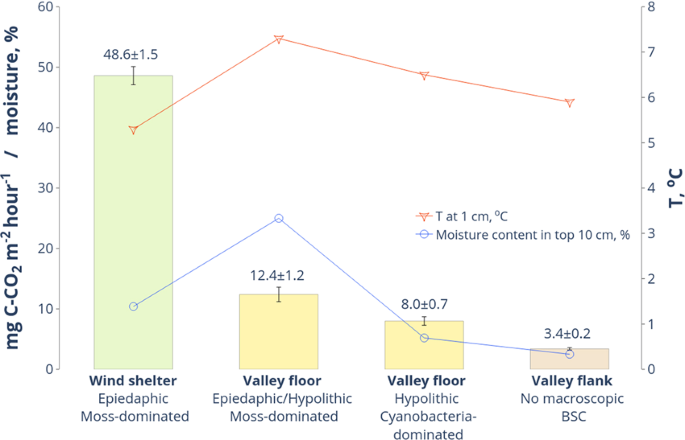
In situ measurements of CO2 emission (± standard error of the mean) from the surface of soils with various types of organogenous horizons and completely without them.
“Microbial landscapes” of hypolithic horizons
The field experiment revealed that microbial hypoliths had colonized intact mineral surfaces over the six-year period. After excavation of glass slides, we observed the “microbial landscapes” that formed in their natural hypolithic environment and consisted of cyanobacteria, unicellular green algae, diatoms, unicellular heterotrophic prokaryotes and fungi (Fig. 8). Colonization replicated the hypolithic pattern with a maximum of microbial biomass attached to the central part of the 75 × 25 mm slides, but not to their upper part, located in vicinity to the day surface. Filamentous cyanobacteria and unicellular green algae prevailed among the primary producers (Fig. 8a–e). Clusters of diatoms have also been detected (Fig. 8f) and can contribute to the primary production as well. However, the established hypolithic complex was dominated by the eurybiontic cyanobacteria (often inhabiting epilithic niche) of those genera, which representatives are also known as lichen photobionts, e.g., Calothrix, Scytonema, Nostoc, Gloeocapsa, Stigonema, or simply eurybiontic genera, such as Phormidium. Cyanobacteria cells often had brown and dark red colors, indicating UV-protecting pigments. Cyanobacteria were predominantly filamentous and produced heterocysts (Fig. 8b) facilitating nitrogen fixation during the polar day and raising its availability in soils. Heterotrophic bacteria and fungi were mainly confined to the loci of primary producers with various forms of their interaction, including parasitism on cyanobacteria, germination of mycelium from lichen soredia, association with algae and cyanobacteria (Figs. 8d,g2). Although the fungal mycelium and spores were abundant as also reported in previous study on soils of the Larsemann Hills46 the morphological diversity of fungi was very low with only a few spore morphotypes observed. Specific features included the monodomination of a single fungal morphotype (Fig. 8g1) and the absence of mycelial Actinobacteria (actinomycetes). However, we cannot completely exclude the presence of unicellular Actinobacteria among the clusters of heterotrophic bacteria that were observed on the slides. In the Larsemann Hills, Actinobacteria are the dominant bacterial taxa in soils44 and are also found within algae- and cyanobacteria-dominated layers under stone pavements45. It is likely that heterotrophs follow the patterns initially created on slides by the primary producers. While the six-year experiment documented intensive colonization of the slides by cyanobacteria and green algae, the heterotrophic colonization may take longer. For instance, Actinobacteria in soils of the Larsemann Hills demonstrate significantly higher relative abundances with the longer distance from the glacier44, thus longer periods of soil exposure.
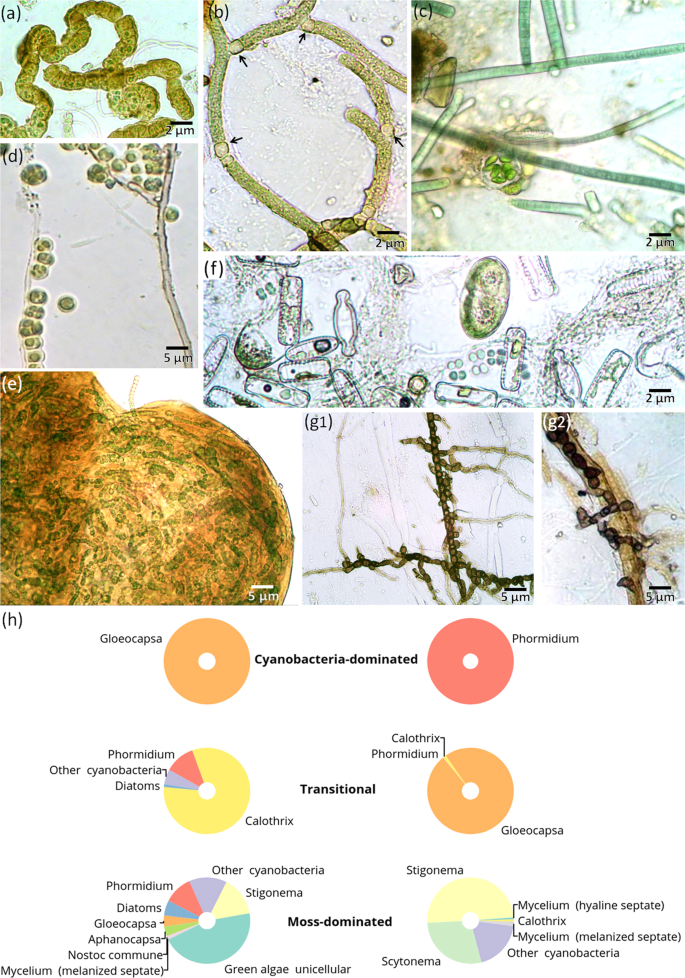
“Microbial landscapes” established on quartz slides after six years of exposure in the hypolithic environment: (a) microcolony of cyanobacteria Stigonema sp.; (b) filamentous cyanobacteria Scytonema sp. with nitrogen-fixing heterocysts (black arrows); (c) spherical microcolony of coccoid Chlorophyta in patchy EPS matrix against the background of Phormidium sp. filaments; (d) unicellular coccoid algae (Chlorophyta) growing along melanized septated fungal hyphae; (e) spherical microcolony of cyanobacteria Nostoc sp.; (f) algae and cyanobacteria association: Diatomeae, Chlorophyta and small coccoid cells of cyanobacteria; (g1) monodomination of the one melanized fungi morphotype with chlamydospore formation and its close association with cyanobacteria (g2); (h) biomass ratios of various microorganisms that colonized slides during six years of exposure in different types of hypolithic horizons.
Surprisingly the slides were not covered by continuous biofilms that are common on subaerial surfaces including those in extreme environments. Instead of continuous coatings the main biofilm-forming agents (cyanobacteria and green algae) formed discrete and compact spherical microcolonies with abundant EPS (Fig. 8e).
The dry microbial biomass constituted 2.4 ± 0.7 mg per cm2 of the slide surface exposed in the cyanobacteria-dominated horizons, 2.3 ± 0.9 mg per cm2 in the moss-dominated and 16.3 ± 17.5 mg per cm2 in the transitional series. The biomass ratios of various microorganisms that colonized slides were discriminated between cyanobacteria-, moss-dominated and transitional series (Fig. 8h), probably inheriting the structure of microbial communities that was already in place. This is in correspondence to the earlier findings20,28,35 on the deterministic processes and successional patterns in hypolithons indicating cyanobacteria-dominated communities as the basal stage. The cyanobacterial hypolithic horizons of the high center polygons produced monodominant pattern of colonization with either Gloeocapsa or Phormidium as the only taxa developing in attached state on the slide surface. The cyanobacterial diversity increased and diatoms appeared in the transitional hypolithic series, while the highest diversity of cyanobacterial and algal autotrophs, as well as pronounced hyaline and melanized mycelium manifested on slides from the moss-dominated hypolithic horizons.
Thus, the six-year field experiment has shown that the new mineral surfaces are rapidly colonized by the microbial autotrophs within the few years of exposure in hypolithic environment. At this timescale the microbial biomass of colonizers is discriminated upon existing microbial patterns in cyanobacteria-, moss-dominated and transitional series of hypolithic horizons, however, continuous EPS-rich coatings known of subaerial biofilms65 do not yet appear.
Hypolithic horizon at a landscape level
In the absence of vascular plant cover, the BSC colonization pattern is clearly recognized on the high-resolution orthoimages in the visible spectrum (Fig. S4) and demonstrates strong connectivity to the soil moisture gradients upon meso- and microtopography of the valleys (Fig. 9 and Fig. S5). We were able to distinguish six classes of BSC on the orthophotomaps, which helped to extrapolate our field data to a landscape level. Hypolithic horizons occurred within three out of six classes of BSC (Table 2): hypolithic moss-dominated + epiedaphic moss-dominated (class III), hypolithic cyanobacteria-dominated + hypolithic moss-dominated with patchy proliferation of epiedaphic moss (class IV), hypolithic cyanobacteria-dominated (class V). The stand-alone hypolithic cyanobacteria-dominated BSC (class V) was present in a 0.5–2 m range of elevation over the talweg (Fig. 9c and Fig. S5c) and was emplaced on the valley slopes (3°) as a 2–20 m wide stripe between the lower-slope mixed moss/cyanobacteria classes (slopes < 3°) and the upper-slope barren ahumic soils (slopes 12–20°). It was not possible to discriminate separately hypolithic BSC in the complex patterns of class III and class IV since the hypolithic BSC often occurred at a very close distance to the epiedaphic one. Such patterns could be reliably resolved only upon a detailed sampling as demonstrated previously at the 10 × 10 m hypolithic site in the valley 1. As far as we learned from the field, the barren ahumic soils can occasionally be enhanced by sporadic inclusions of ephemeral hypolithic cyanobacteria-dominated biofilms, although present only on the undersides of the pavement components. This pattern probably originates from colonization after snowfalls when moisture availability shortly rises. However, it could not be spatially resolved by the techniques we used, and therefore was not accounted in further calculations. When all three classes of BSC that contain hypolithic component are combined, they account for 12.0% of the total soil area at the valley 1 and 49.5% of the total soil area at the valley 2, which is well supplied by meltwater and is one of the most fertile across the Larsemann Hills. Cyanobacteria-dominated morphotype of hypolithic horizon occupied 3.0% and 13.2% of the total soil area in the valley 1 and valley 2, respectively. The other two morphotypes of hypolithic horizons (moss-dominated and transitional) could also be attributed to the BSC classes obtained upon classification of UAV orthoimages. Hypolithic moss-dominated morphotype was present in class III and transitional series of hypolithic horizon were most common in class IV (Table 2). We attributed parameters of the three morphotypes of hypolithic horizons to the three most relevant BSC classes to get a rough upper-limit estimate of how much TOC could be stored by various hypolithic horizons across the soils of Valley 1 and Valley 2 (Table 3). Assessment of TOC densities in a triplet of moss-dominated/transitional/cyanobacteria-dominated morphotypes indicated quite an expected decrease of values from the moss- to cyanobacteria-dominated varieties: 429/86/40 g C m−2, respectively. Taking into account the maximum areas of corresponding BSC classes, the TOC reservoirs for the moss-dominated/transitional/cyanobacteria-dominated morphotypes comprised 2269/361/136 Kg C, respectively. Approximated TOC reservoir of all horizons formed with a major contribution from hypolithic BSC constituted 2765 Kg C at a 36418 m2 of the total soil area in the two valleys. Thus, hypolithic BSC plays at least a distinguishable role in shaping the topsoil and its carbon reservoir at a landscape level, and creates a hypolithic “gateway” for organic carbon to enter depleted soils of the Larsemann Hills.
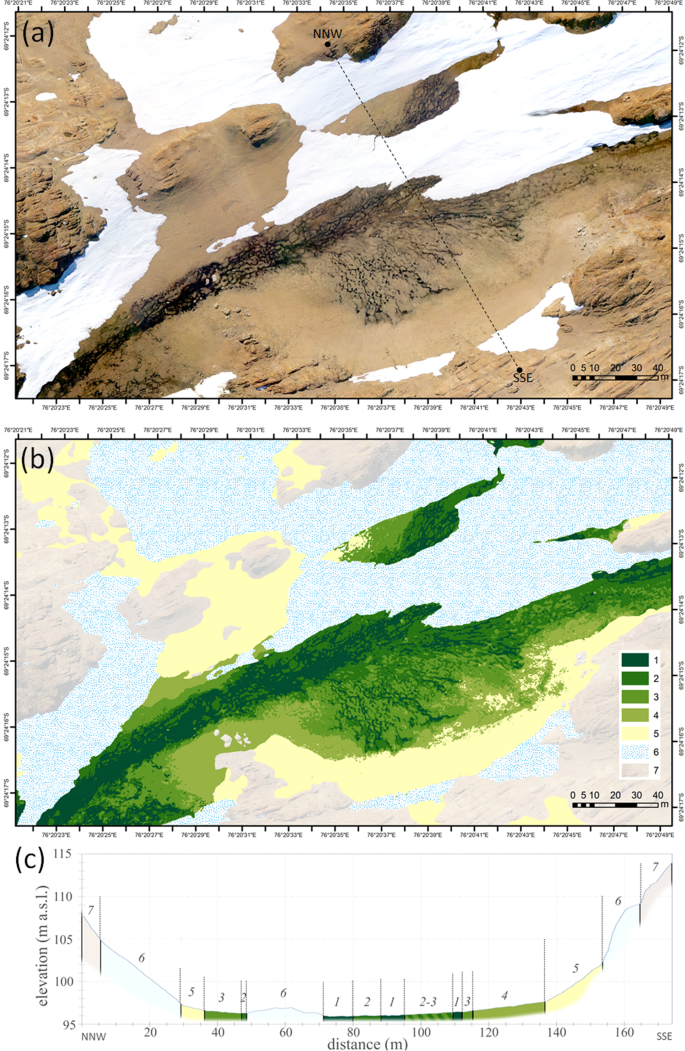
Distribution of BSC in the valley 2 (69.4042°S, 76.3431°E; Larsemann Hills): (a) orthophotomap obtained from mosaic of UAV images as of January 29, 2017; (b) distribution of BSC classes and other landscape components: 1 – amphibian algae- and cyanobacteria-dominated + epiedaphic moss-dominated (class II); 2 – hypolithic moss-dominated + epiedaphic moss-dominated (class III); 3 – hypolithic cyanobacteria-dominated + hypolithic moss-dominated with patchy proliferation of epiedaphic moss (class IV); 4 – hypolithic cyanobacteria-dominated (class V); 5 – no BSC (barren ahumic soil) or ephemeral BSC (class VI); 6 – snow patches; 7 – consolidated rock exposures and rock debris with epi/endolithic colonization; (c) DEM-derived geomorphological profile through the valley as indicated by dashed line on the orthophotomap, the colors and numerals match the legend on (b).
Should the substrate with hypolithic horizon be recognized as a soil
“Hypolithic soils” have been previously recognized in the Atacama Desert by Warren-Rhodes et al.27. Although this denomination was in quotes, it related to the tiny soil layer adherent to the hypolithic communities dominated by cyanobacteria. The current major classifications stay indifferent to soils with cryptic organogenous horizons. Most of the soils in the Larsemann Hills including those with hypolithic horizons have shallow permafrost or bedrock, thus, are classified as Cryosols or Leptosols in the World Reference Base (WRB) system73 and Gelisols or Entisols in the Soil Taxonomy system74. The common soil taxa of the valley floors most conductive to the hypolithic colonization are Protic Turbic Cryosols (WRB) or Typic Haploturbels (Soil Taxonomy). However, hypolithic horizons accumulate a measurable amount of organic matter and form a continuous subsurface body as documented in this study. Moreover, hypolithic horizon is located directly inside the soil matrix and, paradoxically, this fact justifies it as a soil horizon in a more distinct way than the epiedaphic moss-lichen litter, which relation to the soil body is sometimes questioned. The presence of hypolithic hotspot of productivity makes the topsoil enriched in biogenic elements and in general raises complexity of the soil cover in depleted environments of East Antarctica.
There is a set of unresolved questions on the role of hypolithic communities in shaping the soil morphology, aggregates formation (when hypoliths extend to the sandy bedding of the pavement), biomineral interactions, elements cycles and long-term stabilization of organic carbon. It is the future task to explore spatial patterns of hypolithic horizons across the ice-free areas of East Antarctica using such proxy as hyperspectral and multispectral UAV imaging combined with the field sampling. Accurate experiments that could document a link to the deeper soil horizons have yet to be established. Hypoliths are highly dependent on the moisture content and their migrational ability is one of the specific features to explore since some cyanobacteria can actively move to the soil surface or retreat to the refuge below in response to the wetting/drying events75. The contribution of the hypolith-derived organic matter to the soil and landscapes reservoirs should be quantitatively evaluated and related to the other known sources. Having received answers to these questions, we can better understand whether the phenomenon of hypolithic soils is justified and appropriate proposals for soil classification are required.
Source: Ecology - nature.com

