Construction and maintenance of RFs
Construction of RFs is described in detail by Comptour et al.11 and summarized here. RFs in Mossaka are constructed in the floodplain on heavy clay soils where two tributary rivers, the Sangha and the Likouala-Mossaka, join the Congo River (see SI for detailed description of the study area). RFs in Mossaka are permanent structures that, once constructed, are repaired and maintained for decades. They comprise three components, named in the local Likouba language lipu (earth clumps along with rooted vegetation), puleke (topsoil with vegetation debris) and tse (topsoil), which are sequentially piled up on each other (Fig. 2A). RF construction in Mossaka is a (bio-)mass-transfer system: vegetation and soil added to the RF are gathered from the area around the RF, hereinafter termed the RF supply area (RFSA) (Fig. 2B). The total area (RF plus RFSA) usually is about 2.5 times the size of the RF itself.
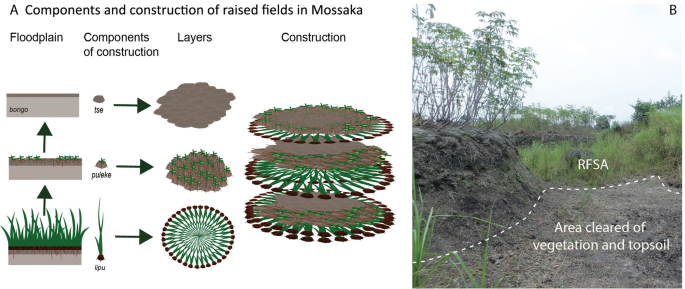
Mode of construction of RFs in Mossaka. (A) Materials used to construct RFs, and sequence of placement. Lipu: earth clumps with rooted vegetation; puleke: topsoil with vegetation debris; tse: topsoil from the RFSA; bongo: grey clay (C horizon) (Ilustration made with Adobe Illustrator). (B) Area cleared of vegetation and topsoil (tse) next to field N2 (which was planted with manioc). Photo (1–20 August 2017), Leonor Rodrigues.
Vegetation used to construct RFs serves as organic amendment. Earth bound by grass roots also provides physical support, allowing the building of mounds high enough (up to 1.5 m) to protect crops from flooding. RFs are mainly planted with manioc, but sweet potato, pineapple, sugar cane and banana are also grown on RFs. Oil palm trees are generally found on abandoned RFs. Over time, much of the plant biomass added to RFs decomposes and after one or two years of cultivation, fields are left fallow for 5–10 years, depending on the farmer. In this biomass-transfer system, fallow periods serve to regenerate topsoil and vegetation not only on the RF but also and most importantly in the RFSA. The RFSA is the property of the field owner, who alone has the right to collect its topsoil and vegetation. When fallow fields are again put into cultivation, new layers of all three components (lipu, puleke and tse) are collected from the RFSA and added to the hoed RF surface.
Physical characteristics of soil profiles in RFs
Soil profiles of four RFs (N1–N3 and AF) located in the floodplain of Mossaka (Fig. 1) were studied. These four fields vary in age and represent different phases in the cycle of cultivation (Figs. 3,4). Fields N1 and N2 were actively cultivated, whereas fields N3 (fallow) and AF (abandoned; Fig. 4) were not. The profiles reveal how the mode of construction and maintenance of RFs influences soil vertical organization, and how this organization changes over time. Field N1 was constructed 15–20 years ago and had been left fallow for at least 10 years just until two weeks before our study, when it was rehabilitated after addition of 50 cm of topsoil and vegetation from the RF and RFSA. Field N2, first constructed more than 40 years ago, had been in fallow for about nine years when it was rehabilitated in 2016 and planted with manioc. This field was thus in its second year of cultivation when the trench was opened (tubers are harvested one to two years after planting). Field N3, built 15–20 years ago, was in fallow when we opened the trench, and had lain fallow for about 10 years. The age of field AF is unknown, but according to local inhabitants, it has been abandoned for more than 20 years. It is thus older than fields N1 and N3.
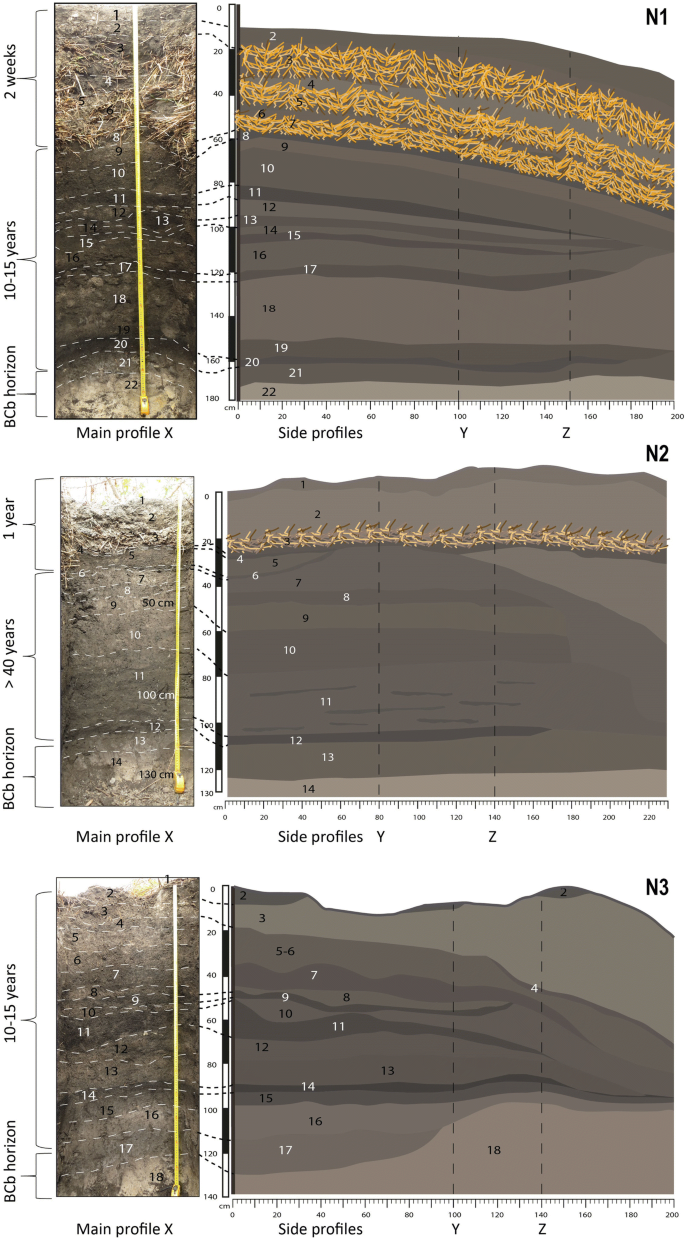
Age and internal morphology of RFs N1–N3. Layers are labeled with numbers. Left panel for each field presents the main profile (X), right panel the two side profiles (Y and Z). Note: layer 1 in side profiles of RF N3 is missing. It was unintentionally removed during our observation and sampling campaign when the field was cleared of fallow vegetation before excavation of the trench. Photos (1–20 August 2017), Leonor Rodrigues and Tobias Sprafke. (Ilustrations made with Adobe Illustrator).
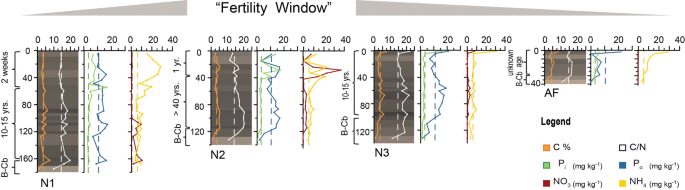
Fertility over time. The three different phases in the cycle of cultivation represented by fields N1 (two weeks after addition of fresh biomass to rehabilitate a field that had been left fallow for at least 10 years), N2 (one year after rehabilitation, field age > 40 years), and N3 (fallow, since about 10 years). Some fields are not only left fallow, but abandoned (AF, abandoned at least 20 years ago). Peak of chemical fertility in the layers where crop roots are concentrated is reached some months to one year after rehabilitation for cultivation (“Fertility Window”). Fertility then declines, returning to values typical of nearby uncultivated soils during the fallow period and falling even lower with abandonment. Plots show downprofile colour (visualized by conversion of spectrophotometrically measured colour space values into the RGB colour space39 and chemical analysis of the main profiles (X) of fields N1, N2, and N3. Dashed lines in the background are the mean values for each chemical parameter of the topsoil (Ah) samples taken at 10 cm depth from areas near each field in the RFSA. (Graphs were plotted with Grapher and arranged using Adobe Illustrator).
As a consequence of the mode of construction, near-surface portions of RF profiles show a sequence of alternating layers (grass/topsoil/grass), conspicuous in the upper 50 cm of N1 (Fig. 3A) and still visible in the upper 30 cm of N2 (Fig. 3B), where layers were applied one year before our study. Within two years after construction, the grass layers are largely decomposed. Below the most recently added layer of vegetation, this results in a characteristic stratigraphy, with dark crumbly layers rich in OM alternating with lighter-coloured clay-rich layers relatively poor in OM. In the oldest fields (N2 and AF), this stratigraphy is much less visible, layer boundaries being much more diffuse than in profiles of fields N1 and N3 (Figs. 3,4). Colour measurements by diffuse reflectance spectroscopy visualized as RGB values in Fig. 439 quantify these field observations and reveal only very weak stratigraphy for N2 and AF. In addition, colour data predict the soil carbon content: across all samples (n = 59), the Lightness value L is negatively correlated with soil organic carbon (Corg) content (R = -0.70, p = 3.3 e−11; Fig. S1, Table S1). The relationship appears to be asymptotic: beyond a certain Corg amount the samples cannot get any blacker (Fig. S1 and Table S1). A well-defined 5–10 cm-thick, black, OM-rich layer with sharp boundaries developed at the bottom of each RF (Figs. 3,4). It resulted from decomposition of the first layer of vegetation deposited during the field’s initial construction (first circle of lipu, i.e. rooted vegetation with earth clumps; Fig. 2). Beneath this layer occurred the natural BCb horizon, composed of yellow-white clay (Fig. 3). The ancient topsoil (Ahb) of the floodplain soils is missing at the base of each RF, as it was removed and incorporated into the first layer of lipu.
Chemical characteristics and structure of soil profiles
The clay-dominated sediments of the floodplain are extremely weathered, containing almost no bases and consisting largely of silica (mean 59%) and aluminum (mean 20%), resulting in a very high CIA (chemical index of alteration40 of 95 (Table S2)). These results are consistent with the measured geochemistry of rivers of the Congo Basin, which are among the most acidic rivers of the world, with low solute concentrations and low amounts of suspended sediments, mainly kaolinite41. Thus, as expected, pH values of all soil samples were very low (mean 3.58, standard deviation ± 0.09). In all samples, Corg content was generally high (mean value 36 g kg−1), ranging from 7.5 g kg−1 in the OM-poor pale clay layers at the bottom of the profiles up to 90 g kg−1 in some OM-rich black layers (main profiles: Fig. 4 and Table S3; all profiles: Table S3). Values of C/N were generally high (mean = 16) and became slightly larger towards the bottom (max = 21.8) of the profile in all fields, pointing to hampered C mineralization and possible biomass-derived N leaching during the anoxic conditions that occur during annual flooding. The reference samples taken from areas adjacent to RFs had minimum and maximum Corg values of 27 g kg−1 and 48 g kg−1, respectively. The oldest field (N2) had slightly lower Corg values than the three other fields and the differences in values between layers were much smaller (Fig. 4; Table S3).
Average total phosphorus (P) content from clay underlying the fields was 400 mg P kg−1 (Table S2), which is moderate compared to usual global ranges in unmanaged soils (< 100—1,000 mg P kg−1) 42. Usually, in acidic soils less than 1% of total P is available to plants, as P is bound to elements such as Al and Fe in insoluble complexes43. Strikingly, we measured relatively high values of total extractable phosphorus Ptex (sum of organic phosphorus (Po) and inorganic phosphorus (Pi)) in the OM-rich layers of the RFs (mean 16.0 mg kg−1, max. 49.4 mg kg−1) and the reference soils (mean 10.3 mg kg−1, max 21.9 mg kg−1) (Fig. 5 and Table S3). Given the absence of chemical fertilizer applications and low atmospheric input42, local biochemical processes seem to make P available. Assimilation of P from local sediment by grasses adapted to anaerobic conditions is likely possible during the flooding period, when P is released upon reduction of Fe3+ to Fe2+44. Crops require inorganic P (Pi) 45, which shows a striking predominance in the upper part of RFs N1-N3, whereas in the lower parts of these fields, in the abandoned field (AF), and in the reference soils Po predominates. Pi becomes available several months to a few years after organic amendments, as maximum contents are in recent former surfaces (which were in fallow before the application of the new material) in N1 (main profile X 50–57 cm) and N2 (profile X 28–29 cm; 37–40 cm). In N2, these layers with high Pi values derive from vegetation applied one year before our observations and sample collection. In the fallow field N3, high Pi values are restricted to a thin surface layer (main profile X 0–5 cm). In the abandoned field (AF) Pi values are low throughout the profile, even in the surface layer.
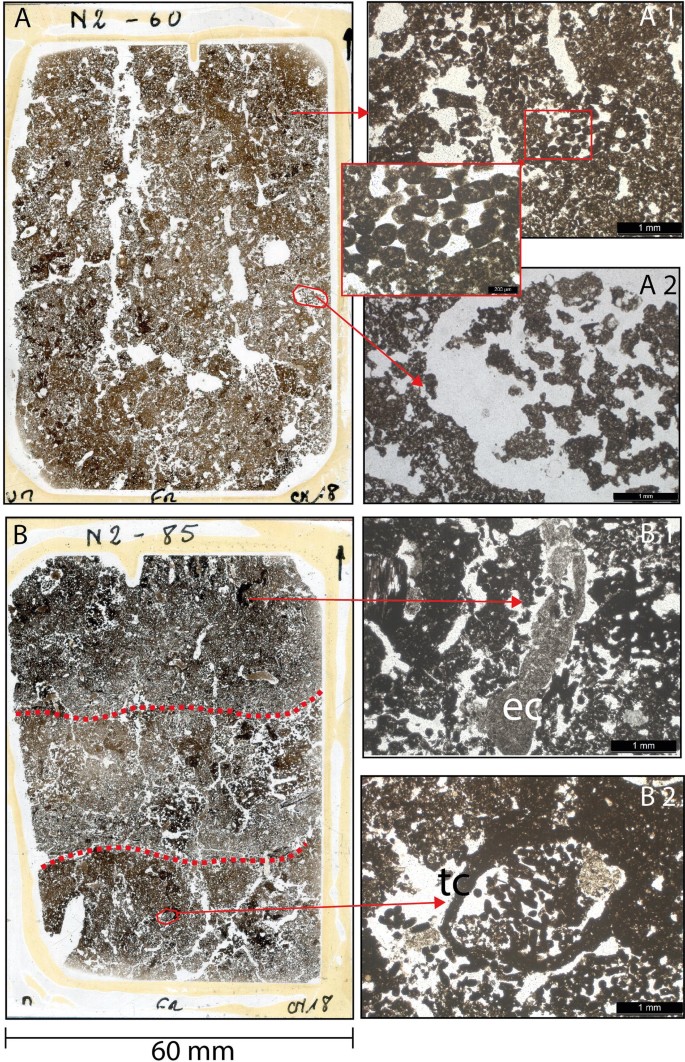
Scans and photomicrographs [plain polarized light (PPL)] from thin sections of field N2 illustrating the alteration of RF soils by biota. (A) Thin section from 60 cm depth, a mixed organo-mineral layer with large bio-pores and root channels infilled with faunal excrements. (A1) Soil structure made from bacillo-cylindric faunal excrements. (A2) Soil pore filled with degraded faunal excrements. B: Thin section from 85 cm depth encompassing three distinct layers (organic-mineral-organic) with boundaries (dashed red lines) blurred by mixing. (B1) Earthworm excrements (ec). (B2) Channel built by termites/ants (tc) infilled with bacillo-cylindric faunal excrements, most probably produced by small annelids (Enchytraeidae) or mites. Thin sections were analyzed with a Leica DMLP 2000 microscope. For complete documentation of micromorphology, see SI.
The results for mineral nitrogen (N) follow a similar pattern. Mineralization under the very acid conditions observed in our study site leads to high ammonium (NH4+) values in the upper layers of the fields (Table S3). Nitrification, like P availability, is normally hampered in low-pH milieus, and values of nitrate (NO3–) are very low throughout profiles of fields N1, N3, and AF. However, nitrate is present in relatively high amounts in field N2, but only in the uppermost actively decomposing layers of the field (highest value 37.3 mg kg−1, found at 30 cm). Recent studies demonstrate that nitrification can occur in very acid soils when sufficient substrate is present46. The high microbial activity in the uppermost layers of the field could induce release of both nitrate and Pi from organic forms remaining in the biomass. Organic matter, crop and microorganisms must interact in some way to produce these high values of Pi and NO3–. Whatever the mechanisms involved, they appear to come into play only after considerable mineralization of OM has occurred, i.e., one year of cultivation of the fields (cf. values for the upper layers of field N2 with those of field N1, to which vegetation was added only recently).
Soil fauna, bioturbation and micromorphology
Field and microscopic observations indicate the abundance of meso- and macrofauna in the studied RFs. Fallow fields (including field N3) frequently harbor colonies of social insects (termites and ants; see SI for detailed information). Profiles of RF soils and examination of soil micromorphology in thin sections (Fig. 5) show the importance of biota in making RF soils more favorable for crop growth. The method of construction of RFs, with relatively loose vegetation layers sandwiched between topsoil layers, creates a porous structure, and porosity is further increased by activity of soil biota, such as extensive channel building by earthworms and plant roots (see SI Fig. S2 and S3, and Table S4). RF soil is crumbly in structure, with abundant biogenic aggregates. In the upper layers of RF profiles, pores accounted for up to 26% of the surface area of thin sections. This was more than three times the value found in the bottom horizons (7%), which correspond to the ancient natural soil horizon (BCb) (Table S4). This enhanced soil structure is particularly important for the growing of root crops such as manioc, whose tuberous roots must displace soil in order to grow.
Most remarkably, soils of Mossaka RFs are completely reworked by soil mesofauna. Thin sections from OM-rich layers (Fig. 5) show abundant dense concentrations of fecal pellets of unidentified soil mesofauna (probably mites, enchytraeids or collembola). Together, soil macro- and mesofauna and plant roots create new habitats for microbiota, disperse them with their excrements (macrofauna) and facilitate microbial decomposition of organic matter by exudation (roots) and by shredding organic residues before mineralization (macro- and mesofauna)47. Amendments with plant biomass and topsoil, and the decomposition of the added organic matter by abundant and intensively active soil biota, thus appear crucial to the functioning, the fertility and the agronomic performance of Mossaka RFs.
Fate of RF soil profiles over time
The mode of construction and maintenance of RFs produces a characteristic stratigraphy of soil profiles. However, as shown for the two oldest fields, N2 and AF (Fig. 3), most stratigraphic features are quickly lost, and with them the information they provide about soil management practices. The cause is the decomposition and mineralization of the added OM and the intense bioturbation by soil fauna that mixes layers, homogenizing the profile to a depth of at least 100 cm, as seen in the profile X of N2. Below this depth, mineralization and bioturbation are both hampered by the prolonged anoxic conditions due to flooding, as shown by high C/N values and the decreasing amount of biogenic features in thin sections taken from these layers. This leads to preservation of OM from the initial layer of vegetation deposited during field construction, marking the boundary between natural and anthropogenic soil layers.
Many authors have noted the negative impact of bioturbation on the stratigraphic integrity of archaeological sites48. Bioturbation and decomposition both occur rapidly in lowland tropical environments when drainage is adequate and soils are well-aerated. The investigated fields show clearly how rapidly this occurs. Within 40 years, stratigraphy in Mossaka RFs is almost completely lost and with it the history of construction, fallow periods and reuse of RFs. Other important factors contributing to the loss of information are the compaction and erosion of the fields after abandonment. RFs in Mossaka are normally built high enough to escape seasonal flooding. Both compaction of fields, because of mineralization of the organic material and reduced macroporosity of the remaining soil, and erosion, due to heavy rains and flooding, might be responsible for the reduction in height of RFs after abandonment. The height of fields varies according to the local flooding height11. AF is currently only 0.4 m high. By comparing with fields under cultivation next to AF, which are all about 1–1.3 m in height, we estimate that at least 0.5 m of AF has been lost, to erosion of the upper layers of AF, to compaction, or both. Erosion and compaction have also been observed for pre-Columbian RFs26.
These results suggest that if pre-Columbian RFs were constructed and managed in a similar way, these traces would have long since disappeared in the centuries since abandonment. The only stratigraphic trace of management practices observed in Mossaka RFs that might plausibly be expected to persist for centuries (if pedological conditions stay the same) is the OM-rich vestige of the initial layer of vegetation found in the frequently waterlogged soils at the very bottom of present-day RFs, a layer that marks the time of construction.
No accumulation over time of nutrients added to RFs
Other potential signatures of past organic matter amendment practices might be expected to be more persistent than stratigraphy. Long-term human use of soils, including cultivation, can result in enrichment in certain elements, particularly P49, which under acidic conditions is normally stabilized in soil by the formation of insoluble complexes with Fe and Al. Thus P is expected to accumulate in soils amended with organic material49 and is often used as a signature of past human land use. The high levels of Ptex in the uppermost OM-rich layers of Mossaka RF (containing large amounts of partially decomposed grasses) show that the two main grasses used to construct RFs, likinga (Phacelurus gabonensis [Steud.] Clayton,) and litsie (Hyparrhenia diplandra [Hack.] Stapf)11, must be efficient in obtaining P from the mineral soil. Despite these repeated inputs, there is no clear accumulation of P in RF soils. The similar absence of P accumulation in vestiges of Amazonian pre-Columbian RFs does not necessarily indicate the absence of organic amendment practices. First, cultivation can lead to depletion of nutrients in soils (including P), not only by decomposition and erosion but also (perhaps even primarily) as a result of removal with harvested crops. Second, depletion of P is particularly likely where OM applied to fields is exclusively of plant origin, as in Mossaka, because plant biomass contains much lower P concentrations than animal manure44. Thus, any accumulation of P is likely to be modest and counter-balanced by removal through crop harvest. Agricultural systems without livestock and their manure and without mineral fertilizers are generally characterized by P-mining conditions50. The ability to mine P should be particularly great for plants adapted to grow in redoximorphic soils (such as in seasonally flooded savannas), where P is more easily mobilized44. As in some other studies36,49, our results thus suggest that neither Corg nor P are notably accumulated over time in this system, as the values in the oldest fields N2 and AF (with the exception of Corg at the base of RFs) are not considerably higher than those in the reference sites.
Chemical fertility is increased during field cultivation, but rapidly declines in fallow fields (Fig. 4) and is even lower in abandoned fields (e.g. Po) (Fig. 4). After one year of cultivation both Pi and NO3– (the most important nutrients for crops) occur in high concentrations, while in the fallow field N3 (except for the uppermost 1 cm) and the abandoned field AF, values decrease to levels comparable to those in the reference soils (Fig. 4) or even lower (Po in AF). The application of biomass and the resulting activities of soil animals that are important for the increased fertility of RFs also have another effect: over time, they largely efface the evidence of the organic matter amendments on which their activity and abundance, and the fertility of RFs, depend.
Lessons for Amazonian pre-Columbian RFs from this ethnographic analogue
Our characterization of soil profiles of present-day RFs in Africa yields insight into how humans transform soil properties, and how soil properties over depth profiles are transformed through time. The understanding of these “short-term” dynamics is essential for interpreting how soils of similar agricultural systems of the past would be transformed over long periods of time, i.e., centuries or millennia. Our results clearly show that the construction and management of RFs, in combination with bioturbation, result in considerable improvement of soil properties when fields are cultivated compared to the unmanaged natural soils in the surroundings. Our analysis of soil micromorphology shows that strong bioturbation occurs within cultivated RFs, which on the one hand enhances soil structure and nutrient availability, but on the other hand also erases the initial stratigraphy resulting from the field’s initial construction. In the long term, this leads to the loss of information on how fields were constructed and how they functioned when they were cultivated, hampering the interpretation of ancient agricultural systems. Such stratigraphic information may be preserved only under special circumstances, for example, when RFs are buried by fluvial sedimentation, as were Maya RFs in Belize25.
The same features as found in Mossaka—periodic flooding, low nutrient contents of highly weathered soils, grasses adapted to redoximorphic soils where P is more soluble, stockless agricultural systems with no animal manure—characterized the natural and agricultural environment of pre-Columbian RFs in Amazonian savannas9,24, 27,28,29,51. So far, soil composition offers no evidence for any organic amendment practices on pre-Columbian RFs: values for charcoal, SOM and P are not higher on RFs compared to adjacent reference soils13,27. This mirrors our results in Mossaka, where nutrient availability in the fallow field (N3) and the abandoned field (AF) differs little from that in reference topsoils. If only abandoned or fallow RFs had been studied in Mossaka, this would have resulted in great underestimation of their agronomic potential. In contrast, nutrient availability is much higher in the cultivated RFs. Our results suggest that when pre-Columbian RFs were under cultivation, addition of vegetation (with adherent topsoil) as organic amendment could have conferred great agronomic benefits. This practice would not have resulted in accumulation of OM and nutrients in RF soils. As with stratigraphy, evidence of nutrient enrichment would have been erased by the very functioning of the soil system. Absence of evidence for nutrient accumulation thus cannot necessarily be taken to indicate the absence of organic amendment practices.
No present-day subsistence system provides a perfect analogue of a past system. Environments vary; environments influence, but do not determine, cultural adaptations; and human agency allows different solutions to similar problems52. Concerning RF agriculture, as with other kinds of cultural niche construction in seasonally flooded savannas, pre-Columbian systems in Amazonia may resemble present-day African systems in general53 but also differ from them in important aspects54. Within Africa, how RF agriculture is conducted shows important differences between Mossaka and another recently studied site, in the Bangweulu Basin in Zambia10,55. How pre-Columbian Amazonian RFs functioned is likely to show similar variation among sites. Nevertheless, our results from Mossaka show that the conclusion that nutrient-management practices were unimportant in pre-Columbian RFs28, 29 is at the very least premature. Our study joins the few that discuss a fundamental problem in geo-archaeology—to what degree can traces of ancient agricultural practices be stored in soils?37. Our results underline the importance of comparison with present-day analogues in interpreting past agricultural systems and suggest promising lines for future geo-archaeological investigations.
Source: Ecology - nature.com

