Climate change is predicted to dramatically alter precipitation regimes and global hydrological cycles1,2,3. Although changes in the spatial distribution of rainfall can both mitigate and amplify differences between dry and wet regions, there is a consensus that many regions will suffer severe impacts of increased variability and magnitude of precipitation1,2,3,4. These climatic fluctuations can cause extreme hydrological events, such as flooding and drought, which can lead to widespread, though largely unknown, shifts in ecosystem structure and function, particularly in freshwater ecosystems5,6,7,8. Such events can expose some ecosystems to conditions with no recent historical precedent6. Whereas experimental research has focused on incremental changes in mean conditions, fluctuations or extreme events such as floods and droughts may have more profound ecosystem consequences6. In addition, most studies simulating precipitation extremes at the multi-site scale have been limited to a single trophic level, particularly producers5. Therefore, there is a clear and urgent need to identify which food web compartments (decomposers, primary consumers, predators) are most vulnerable to rainfall fluctuations and extremes5,6.
Whereas multitrophic research has focused on a single direction of extreme change, especially drought7,8,9,10,11,12,13,14,15,16, the ecological consequences of drought and flooding have rarely been explored in concert17. Such studies suggest that drought can substantially alter aquatic7,8,9,10,11,12,13,14,15 and terrestrial food webs16,17,18,19, with consequences for community structure and ecosystem function. Drought also weakens trophic cascades and the strength of biological interactions (e.g., competition, predation), and disproportionately threatens top predators, often resulting in communities dominated by smaller organisms6,7,8,9,10,11,12,13. However, previous work has provided limited mechanistic understanding of differences in ecosystem sensitivity to global change5, has been conducted at a local scale6, and has used dissimilar experimental approaches and methods5. This hampers our ability to predict global impacts of drought and flooding on multiple taxa and trophic levels, including standing stock biomass, trophic biomass pyramids20,21,22, and biomass fluxes through the food web23, across large geographic regions.
Standing stock biomass is a common metric in food web research, with trophic structure represented by the distribution of biomass across different trophic levels21,22. These biomass pyramids can summarize changes in complexity and biomass flux through food webs20,21,22. Pyramid size and shape exhibit highly variable patterns across different types of ecosystems worldwide20, but it has been shown that climatic stability can change the shape of biomass pyramids24. Thus, quantifying biomass pyramids improves mechanistic understanding of climate change effects on food web structure, resource partitioning, and energy use.
Here we conducted a geographically coordinated experiment5 in seven sites across Central and South America (18°N to 29°S, Fig. 1a) to investigate the effects of rainfall changes on trophic structure24,25. We used natural, detritus-based microcosms (bromeliad phytotelmata) as model systems due to their widespread distribution and ease of manipulation10,11,12. Bromeliad aquatic ecosystems are inhabited by a diverse fauna10,11,12, comprising top predators, mesopredators, and detritivores15,24,25. We contrasted rainfall-mediated changes in hydrological stability of the study system with the effects of two main rainfall components: (i) the mean daily amount of rainfall, μ; and (ii) distribution of rainfall events around this mean through time, k (i.e., a measure of evenness in the frequency distribution of rainfall; hereafter “frequency”; Fig. 1b). For instance, reductions in both mean daily rainfall (low μ) and the even frequency of rain (low k, hereafter “infrequent rainfall”) increase the proportion of days that bromeliads are empty of water. Regular (current) variability of the rainfall components in each site were first determined using recent meteorological data (see Methods). We applied a negative binomial distribution to these data to estimate the parameters μ and k. We applied ten levels of μ (ranging from 0.1 to 3.0) and three levels of k ranging from 0.5 to 2.0 in a fully factorial experimental design at each of our seven sites for a total of 10 × 3 × 7 = 210 food webs in individual bromeliads. This allowed us to compare ambient, baseline conditions (μ = 1, k = 1) and extreme fluctuations of rainfall quantity (10–300%) and frequency (50–200%) to average historical levels of daily variability for each site (“Methods”).
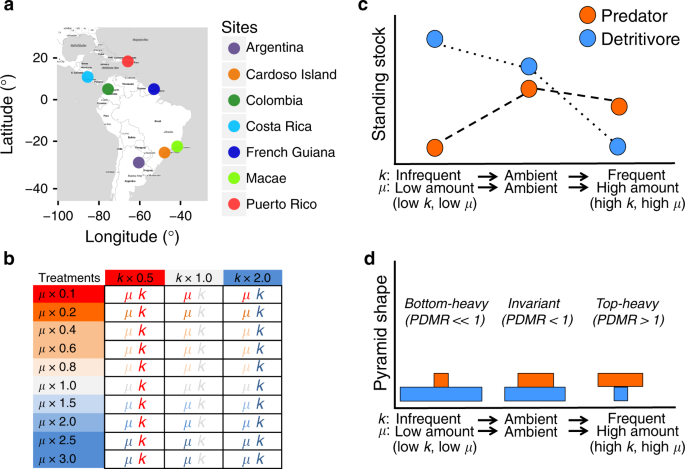
a Study sites. b Treatment combinations designed to manipulate the quantity of rain entering the ecosystem (50–200%) and distribution of rainfall frequency (10–300%) relative to average ambient conditions of each site (μ = 1, k = 1, in the centre of the table). Gradients of rainfall conditions, from severe drought to frequent rainfall, are represented by red and blue colours. k is the dispersion parameter (a measure of evenness in the frequency distribution of rainfall), and μ is the mean parameter (a measure of mean daily rainfall). These parameters were calculated per each site based on rainfall patterns of the last five years before the beginning of each experiment (see “Methods”). Stronger impacts of rainfall events are predicted when their components depart from current scenarios. c We predict that larger predators are more sensitive in environments experiencing infrequent and low rainfall amount (low k, low μ), characterized as extreme drought conditions, than smaller organisms. Under these conditions we expect communities dominated by smaller organisms (detritivores and filter feeders), which tend to decrease in standing stock under more favorable conditions (ambient) via top-down control. In contrast, more frequent rainfall and high rainfall amount (high k, high μ), characterized as heavy rainfall, could impact lower trophic levels through hydrodynamic perturbations (e.g., overflow of nutrients and basal resources). Bigger predators could be more resistant to flooding, but their biomass could decrease slightly via bottom-up effects. d Changes in standing stock, in turn, could alter predator–prey mass ratios (pyramid shape). Drought, ambient and heavy rainfall conditions could cause bottom-heavy, invariant and top-heavy biomass pyramids, respectively.
We hypothesized that precipitation would affect trophic structure in bromeliads by altering hydrology, that is, the temporal dynamics of water within the bromeliad. We therefore also quantified the underlying hydrological dynamics within bromeliads in the field (see “Methods”) as a potential proximate driver, and then projected it in multivariate space using principal component analysis, after standardization between sites (Supplementary Table 2, Supplementary Fig. 1). We defined top predators as species without natural predators within the aquatic food web. All bromeliads were open to colonization and extinction throughout the two-month experimental duration and thus communities could dynamically assemble or disassemble.
The multi-site approach allowed us to explore generalities and site contingencies in food web responses to climate change. We expected stronger impacts of rainfall events when their components departed from current scenarios (μ ≠ 1, k ≠ 1). If bigger predators are more sensitive to drought (here measured as lower values of μ and/or k) than smaller organisms (e.g., mesopredators and detritivores)6,7,8,24, then drought could have stronger ecological impacts than heavy rainfall6. Consequently, under drought or extremely infrequent rainfall events we expected communities dominated by smaller organisms7 (especially detritivores and filter feeders), which tend to be suppressed via top-down control under more favorable conditions (Fig. 1c). Conversely, an excessive amount of rainfall combined with higher frequency of rainfall among consecutive days could impact lower trophic levels through hydrodynamic perturbations, e.g., if important nutrients and basal resources (detrital organic matter and microorganisms) are lost to flooding26,27. Bigger predators could be more resistant to flooding, but their biomass could still decrease if they have fewer resources to support them. Consequently, such changes in standing stock biomass could alter predator–prey biomass ratios (i.e., pyramid shape; Fig. 1d)20,21,24. Because drought and high rainfall frequency are predicted to favor lower and higher trophic levels, respectively, drought is expected to be associated with bottom-heavy biomass pyramids, and high rainfall frequency is expected to be associated with top-heavy biomass pyramids (Fig. 1d). Both the climate and the regional species pool are different among sites12,25,27, which may lead to site-specific contingencies, e.g., communities from arid regions and/or regions with large natural amplitudes in rainfall may be more resistant to drought than those from wet regions and/or regions with narrow climatic amplitudes27.
Extreme precipitation events and underlying hydrological dynamics influence each trophic level in a different manner. Whereas the biomass of detritivores, often the smaller organisms in the study system, is higher under more stable hydrological conditions, the biomass of predators is highest under uneven rainfall (drought conditions). Higher resource concentration under drought conditions fuels these higher trophic levels, resulting in top-heavy biomass pyramids. Our results demonstrate that organisms from lower trophic levels may be the most susceptible to ongoing climate change.
Source: Ecology - nature.com

